(10)日本人のルーツ Ⅰ 常識への挑戦 OWLのひとりごと
![]()
日本人のルーツ(Ⅰ)私たちが聞かされてきたこと
2014.11.7
私たちが聞かされてきたこと 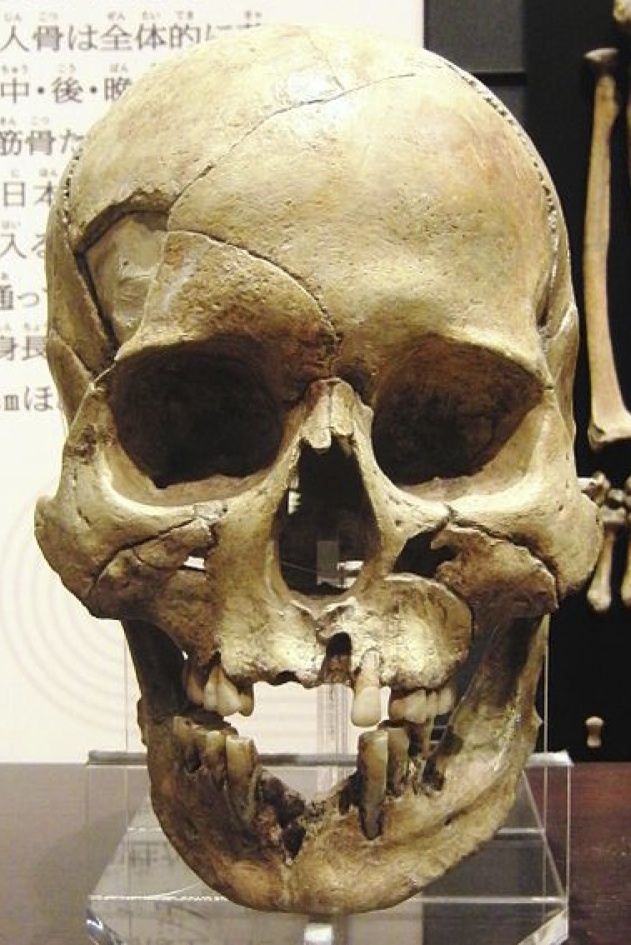

いわゆる縄文人(左)と弥生人(右)の頭蓋骨(女性)
私たちが聞かされてきたこと
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
このページのまとめ
1)日本人は大陸から、樺太、朝鮮、台湾を経由して来た。それが常識だった。
2)誰もその常識に疑問を挟まなかった。
3)形態人類学が絶対視されていた。
4)縄文人が大陸から来た弥生人に置き換わった。そう考えられていた。
5)次の考えは縄文人と北東アジア系の混血で形成されたという二重構造モデルだった。
6)新しい年代測定法や分子遺伝学の成果は、必ずバッシングされた。
7)頑迷な常識が書き換わるには長い時間がかかる。
8)本サイトで常識を疑い、それを書き換えよう。
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
日本人は何処から来たか?
日本人は何処から来たか?この問いに無関心な日本人は少ない。だがその答えはどうか?議論に関心を寄せる人はいない。常識だからだ。
日本人は大陸から来た。樺太経由、朝鮮半島経由、台湾と琉球列島経由で。一部はフィリピンあたりから黒潮に乗って来た。稲作を携えて大陸や朝鮮半島から人々が渡って来て弥生文化を作った。教科書に書いてある。それで終わり。
二〇〇八年の「詳説 日本史の研究」(山川出版社)には、「弥生文化は、農耕社会を形成していた朝鮮半島南部から、稲や金属器をたずさえて日本列島に渡って来た若干の渡来人が、縄文人とともに生み出した文化であると考えられる」と書かれてある。
問題は二つある。一つは「若干の渡来人」、もう一つは「農耕社会を形成していた朝鮮半島南部から、稲や金属器をたずさえて」の部分である。
最初の渡来人の規模に関しては、二〇〇四年の「新編 新しい社会6(上)」(東京書籍)に、「米づくりが広がったころ、朝鮮半島から日本列島へわたってきて住みつく渡来人が大ぜいいました」とある。四年で規模が「大ぜい」から「若干」に改められたようだ。
しかし以前は、渡来系弥生人が縄文人を征服し、縄文人に取って代わったと信じられていた。後述するように、現在この考えは否定されている。いわゆる縄文人から弥生人への変化は徐々に起こった。突然起こったのでも、征服などによって置換されたのでもない。
次は、朝鮮半島が文明の先進地域で、日本列島は最後進地域という部分だ。それが日本人の間で信じ続けられている。
実際はどうだったのか。稲は朝鮮半島から伝わったのか。朝鮮半島南部は本当に農耕社会を形成していたのか。当時、金属器具は朝鮮半島にあって、日本にはなかったのか。その答えとして、次のような事実が明らかになっている。
稲には、その遺伝子配列の特徴により、aからhの八種類がある。シナ大陸には八種類すべてがそろっている。朝鮮半島ではbを除く七種類が存在する。ところが日本列島で発見される稲の種類はaかbのどちらかだ。よって稲作が朝鮮半島経由で伝わった可能性はない。
紀元前十世紀頃、北九州では灌漑設備が整った水田稲作が始まっていた。韓半島で同様な遺跡はまだ発見されていない。農耕社会を形成していたのはどちらだったのか。
鉄器に関しても日本最古の鉄製鋤先はBC三九〇~三四〇年頃と推定されるのに対し、朝鮮半島では紀元前三世紀とされている。日本の方が百年近く早い。
これらから疑問がわいてくる。日本列島が文明の最後進地域だという常識は、真実であったのだろうか、と。
私たちの常識とは、先進文化をたずさえた渡来人が来て、最後進地域である日本に文明の光をもたらしたというものだった。その常識に疑問が投げかけられた理由を、次項以降において、もっと詳しく記すことになる。
しかしその前に、こうした常識はどのように作り出されていったのだろうか。まずはざっと見てみよう。
エドワード-モース
明治期に東京帝大教授として招聘されて来日した米国人動物学者エドワード・モースは、大森貝塚を発見したことで有名だ。彼の主張は次の通りである。
1)本土には本土人ともアイヌ人とも違う縄文人が住んでいた。
2)縄文人は今の日本人の祖先とは言えない。
3)記紀の「国生み」「天孫降臨」「神武東征」などは、
天皇の祖先が渡来し、先住民を征服したことを物語っている。
4)縄文土器を作った人々と弥生土器を作った人々、
すなわち縄文人と弥生人は連続していない。
5)日本人の祖先は縄文人ではなく弥生人である。
この説は大正期以降に定着し、私たちの常識を作った。今もそう信じて、たとえば「天皇家は朝鮮半島がルーツだ」と真顔で言う学者、評論家、政治家、教師もいるほどである。
形態人類学
形態人類学は形質人類学とも自然人類学とも呼ばれる。主に発掘された人類や霊長類の化石を対象に、骨や歯の形態から食生活、運動様式、生活環境、社会構造などを明らかにしようとする。身長、頭長、頭幅、鼻骨の隆起度などから分類してゆくのである。
縄文時代と弥生時代の人骨を並べると、後者は前者と比較して次のように変化している。
a)身長は高くなり、
b)頭は前後に長くなり、
c)鼻が低くなる。
d)縄文人と弥生人の中間に位置する人骨は発見されていない。
こうした比較から、弥生人が縄文人と同じ人種とはとうてい思えない。やはり縄文人から弥生人という別人種に置き換わったに違いない。弥生人とは渡来系の人種だったのだ。そう考えたわけである。
一方、元東大教授の鈴木 尚氏(すずきひさし:昭和三〇年~東大在職)は、縄文時代から現代まで、関東地方で発掘された人骨を形態学的に比較した。その結果から、同じ関東人でありながら、頭骨を含む全身の骨格があたかも別人種ではないかと思えるほど、時代とともに変容したことを明らかにした。
古墳時代から戦国時代までは鼻が低く、縄文時代と江戸時代の人は鼻が高かった。背は古墳時代と明治以降で高く、縄文時代や室町から江戸末期までは低かった。頭の長さも弥生時代から鎌倉時代にかけて長く、縄文時代や室町から現代までは短かった。
弥生人は長身と言われたが、縄文人より五センチ程度伸びたに過ぎない。千年オーダーで五センチである。明治以降百年余りで十センチ以上変化したことに比べて、大した変化とはいえない。
宮家や徳川将軍家の人々の顎は小さく、細面(ほそおもて)だった。一般の日本人と別人種だったわけではない。食べ物によって咀嚼力に違いを生じ、顔面骨格の変化をもたらしたのだ。こうして鈴木氏は、日本人はほぼ同質な集団であると考えた。
ところが鈴木氏の後継者である埴原和郎氏(はにわらかずろう:昭和四七年~東大在職)は、渡来者の数は相当多かったに違いないと推定した。そして日本人形成史の仮説、いわゆる「二重構造モデル」を提唱した。
ただし、根拠と呼べるような論理が埴原氏にあったわけではない。単なる推定、推論だけで結論付け、仮説を提唱した。そうした批判を浴びている一面もある。
二重構造モデルとは、「旧石器時代以来、日本列島に住むようになった東南アジア系の縄文人基層集団の上に、弥生時代以降に渡来した北東アジア系集団が覆いかぶさるように分布して混血することによって現代日本人が形成された」というものだ。
もっとも、基層集団として東南アジア系ではなく、北東アジア系集団を想定する考えも強いようだ。
埴原氏の仮説は、縄文人と弥生人が全くの別集団で、征服されて置き換わったという考えとは違う。異なる二集団の混合によると考えたわけである。しかし二重構造モデルは有力なものとされ、やがて「常識」の一部となった。
このように形態人類学は、日本の学会の中心的役割りを担いつづけ、ごくごく最近まで世の常識を作ってきた。あるいは現在でも、その常識がまかり通っていると言って良い。
司馬遼太郎、山本七平の影響
では、作家や評論家の場合はどうだっただろう。彼らは日本人の常識や世論形成に大きな影響を与えた。
まず作家の司馬遼太郎氏は次のように述べている。
「日本民族はどこからきたのでしょうね」
「しかしこの列島の谷間でボウフラのように湧いて出たのではあるまい」
「我々には可視的な過去がある。それは遺跡によって見ることができる。となれば日本人の血液の中の有力な部分が朝鮮半島を南下して大量に滴り落ちてきたことは紛れもないことである」
「日本人の血液の六割以上は朝鮮半島をつたって来たのではないか」
「九割、いやそれ以上かもしれない」
「ともあれ縄文・弥生文化という可視的な範囲で、我々日本人の祖先の大多数は朝鮮半島から流れ込んできたことは、否定すべくもない」
(司馬遼太郎著『街道を行く1 湖西のみち』週刊朝日、一九七一年)
「まさしく日本列島は、太古以来、文明という光源から見れば、紀元前三〇〇年くらいに、稲を持ったボートピープルがやって来るまで、闇の中にいました。この闇の時代のことを『縄文時代』といいます。旧石器時代に続く時代で、この狩猟採集生活の時代が八千年も続いたというのは、驚くべきことです。文明は、交流によってうまれます。他の文明から影響を受けずにいると、人類は何時までも進歩しないということを雄弁に物語っています」
(司馬遼太郎『文学から見た日本歴史』ケンブリッジ大・英国日本学研究会主催シンポジウム特別講演、一九八九年)
評論家の山本七平氏も次のように語っていた。
「日本人は東アジアの最後進民族です。先進・後進を何によって決めるか、どのような尺度を採用するかは相当に難しい問題でしょうが、例えば数学ですね。中国人は偉大な民族で、西暦紀元ゼロ年頃、既に代数の初歩を解いていたのですが、当時の日本人ときたら、やっと水稲栽培の技術が全国に広がったらしいという段階、まだ自らの文字も持たず、統一国家も形成しておらず、どうやら石器時代から脱却したらしい状態です。
この水稲栽培、即ち農業に不可欠なのが正確な暦ですが、ヨーロッパ人がメトン法(十九年七閏の法)を発見したのが紀元前四三二年、一方中国人は紀元前六〇〇年頃に既にこれを発見していました。中国人は当時の超先進民族です」
(山本七平著『日本人とは何か 上』PHP、一九八九年)
彼らは超有名人である。ファンも多い。司馬遼太郎や山本七平が言うのだから間違いがない。進歩的で知識の最先端を把握していたはずである。こうして私たちの常識は確固たるものになる。
常識ができあがった後
いったん常識化すると、その後は学会でどんな発表がされても、通説に基づいて真偽が判断された。学会とはサムシング・ニューを発見し問いかける場であるはずだ。しかし日本の学会は違う。これまでの学説を支持する発表が「良い研究」とされた。
二〇世紀の終わり頃、新しい年代測定法や分子遺伝学が登場した。すると歴代の学者たちは頑迷に抵抗した。日本の方が古い!と発表すると必ずバッシングされた。二重構造モデルに疑問を投げかけると無視された。素晴らしい研究だと歓迎はされなかった。何故か?
土器の年代、稲作が到来した時期、日本人のルーツ、文化など、これまで蓄積してきた常識が揺さぶられるからだ。新しい年代測定法や分子遺伝学の成果に基づいて通説が書き換えられると、何か困るのだろうか。彼らの権威が崩壊し、面子丸つぶれになるだけだろうに。
こうして、新しい年代測定法や分子遺伝学の成果は、欧米の学会のトレンドからかなり遅れて日本に取り込まれる。学者の権威や面子がつぶれないよう、ゆっくりゆっくり、何ごともなかったかのように、少しずつ書き換えが始まる。
しかし書き換えと言っても、わずかな修正にとどまる。常識の骨格部分は必ず残るようにしてある。パラダイムがすっかり変わってしまうような激動、激変を、この国の学者たちは好まない。
もちろん公教育では、新たな考古学的事実が反映されるのはずっと後になる。なかなか教科書に反映されることはない。日本の学校では教科書の記述を、概ねそのまま受け入れるのが「良い生徒」である。疑問を呈したり、反論したりするのは「出来の悪い生徒」である。
議論する人間は嫌われ、出る杭は打たれるのだ。
教科書に誤りや噓があっても、それに疑問を差し挟まない生徒が学業優秀で、やがて指導的立場に立つ。学者、教師、教科書執筆者、知識人、文化人、作家、評論家、記者、論説委員、役人、政治家になってゆく。
彼らから私たちは聞かされ続けて来た。社会の常識を。その常識を私たちは疑うことをしない。日本人のルーツ?もう知ってるよ。教科書に書いてある。新事実が出てきた?ちょっとでしょ。大筋は変わらないでしょ、と。
このように、たとい事実が社会常識と違うことがわかっても、私たちの頭に巣食う常識が書き換えられるのは、ずっとずっともっと先になるわけである。
日本人は何処から来たか?
私たちの常識を疑い、それを書き変える作業に入っていこう。
日本人のルーツ(Ⅱ)質量分析計による年代測定法の登場
2013.10
質量分析計による年代測定法の登場 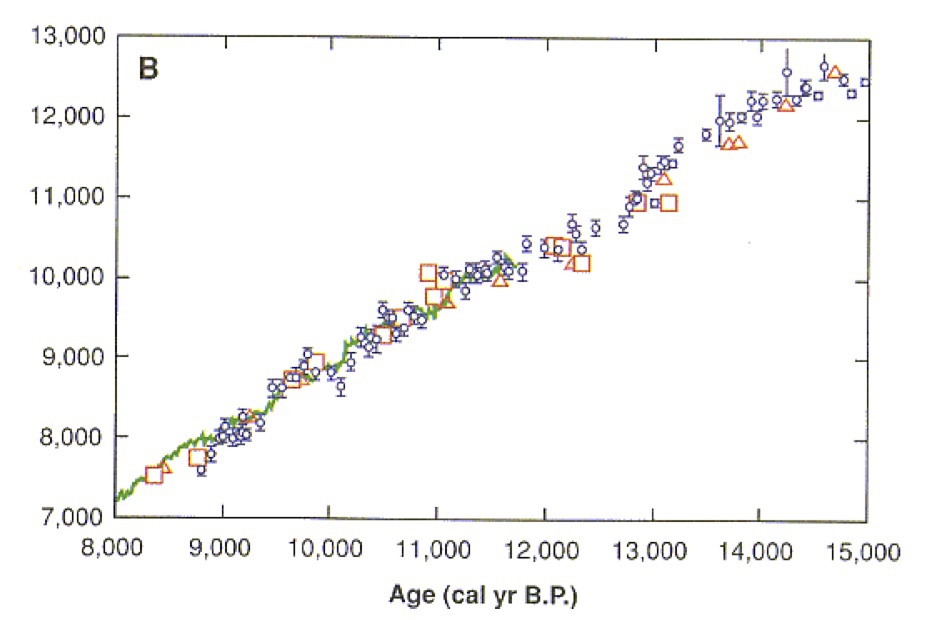
加速型質量分析計によるC14年代推定法の較正曲線
質量分析計による年代測定法の登場

本項では、年代推定法の変遷をたどる。そして加速型質量分析計の登場によって推定暦年の精度が著しく向上したことを述べる。
土器編年法
土器出現の順番は正しかったが実年代は不明だった
日本の考古学の世界でこれまで伝統的に研究されてきたのは、遺跡から出土した土器の種類を細かく分類し、新旧の順に並べること、つまり専門用語で「土器編年」といわれる作業です。
これまでの長い研究の歴史によって、今では相当に細かな土器の編年作業ができています。
こうした研究は戦前の昭和18年ごろに基本的な骨格ができあがり、その後の数十年間でかなり精緻なものになっています。
しかし、土器編年からわかるのは、土器を古い順番に並べることであって、年表にして何年ごろかという具体的な年代ではありません。つまり、相対的な年代であって、絶対年代ではないのです。
たとえば、土器のタイプから紀元前35年とか、紀元後100年というような具体的な年代は絶対に出てきません。ある土器が弥生中期後半のものであることはわかっても、いったい何年から何年ごろまで使われたか、ということは厳密にはわかりません。
あえて年代をつけようとすれば、土器と一緒に出土する遺物などによって、だいたいの年代を推定するだけです。たとえば、土器と一緒に中国の鏡や貨幣が出土した場合は、年代を知る有力な手がかりとなります。それらは中国で造られた時代がほぼわかっているからです。
しかし、その場合でも、中国の鏡や貨幣が海を越えて日本に渡り、遺跡の中に埋もれるまでにどれくらい時間がかかったのか、という問題があります。それは個々の研究者によって、考え方が少しずつ違ってきます。
ですから、出てきた年代はアバウトな時間幅をもち、ある程度の誤差が出てくるのが当然です。多くのデータをそろえ、比較検討して初めて、ある程度正確な年代が推定できるのです。
http://www2.odn.ne.jp/hideorospages/yamatai02.html
C14年代測定法
まず、地球生物圏内では放射性炭素(14C)の存在比率がほぼ一定であると仮定する。すると動植物内部の14C存在比率は、生きている間は一定に保たれる。生物は常に炭素を含む栄養を外界から取り入れ続けるからである。
しかし死後は、もはや新しい炭素が外から補給されなくなる。そのため個体内の14Cの存在比率は下がり始める。
この性質と14Cの半減期が5730年であることを組み合せる。すると新しい炭素が供給されなくなった暦年、すなわち生きていた年代が推測可能となる。この年代測定法により算出された暦年が放射性炭素年代と呼ばれる。
炭素14 (14C) は、約5730年の半減期でβ崩壊をして減じていく性質をもっているため、これを利用して試料中の炭素同位体12/14比から年代を推定することができる。測定限界が元の約1/1000である場合、約6万年前が炭素14法の理論的限界になる(実際の測定では、ベータ線測定法の場合は3 - 4万年程度、AMS法では4 - 5万年程度が測定限界)。
「ガスプロポーショナルカウンティング法」「液体シンチレーションカウンティング法」と呼ばれ、炭素14が電子と反電子ニュートリノを放出して窒素14(14N 普安定同位体の窒素)に壊変するときに放射されるベータ線をシンチレータにより検知して数える方法である。現代の炭素1gでも4 - 5秒に1個しか壊れないので、計測には時間がかかり、試料もグラム単位で必要とされる。
14C → 14N + e- + Ve
ガスプロポーショナルカウンティング法では、炭素14を二酸化炭素のガスに変化させベータ線を計測する。
無機物及び金属では測定が出来ない。
本原理を一九四六年に開発したリビー(W.F.Libby:米国)は、一九六〇年にノーベル化学賞を受賞した。放射性炭素(14C)年代測定法の上限は4~5万年とされている。
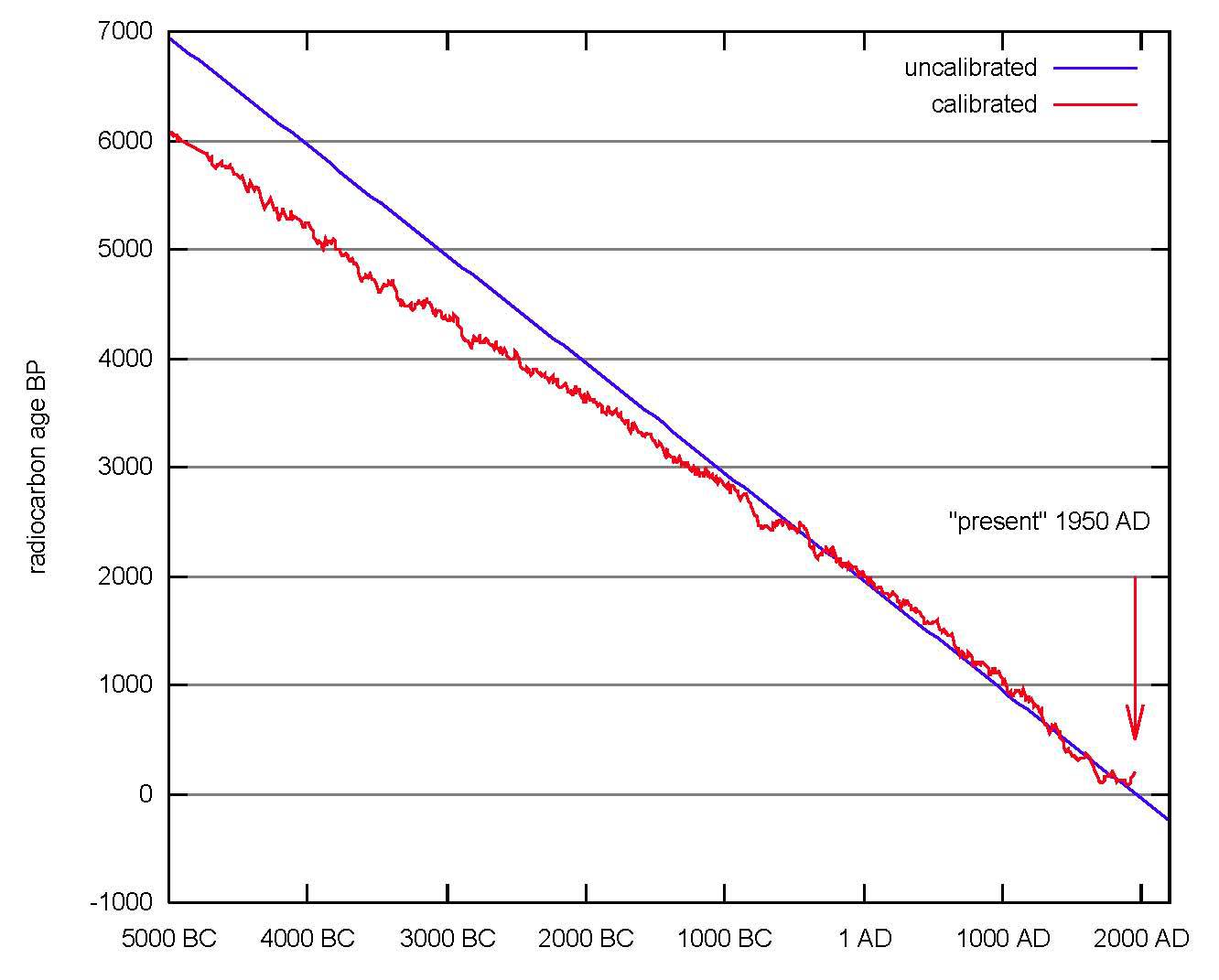
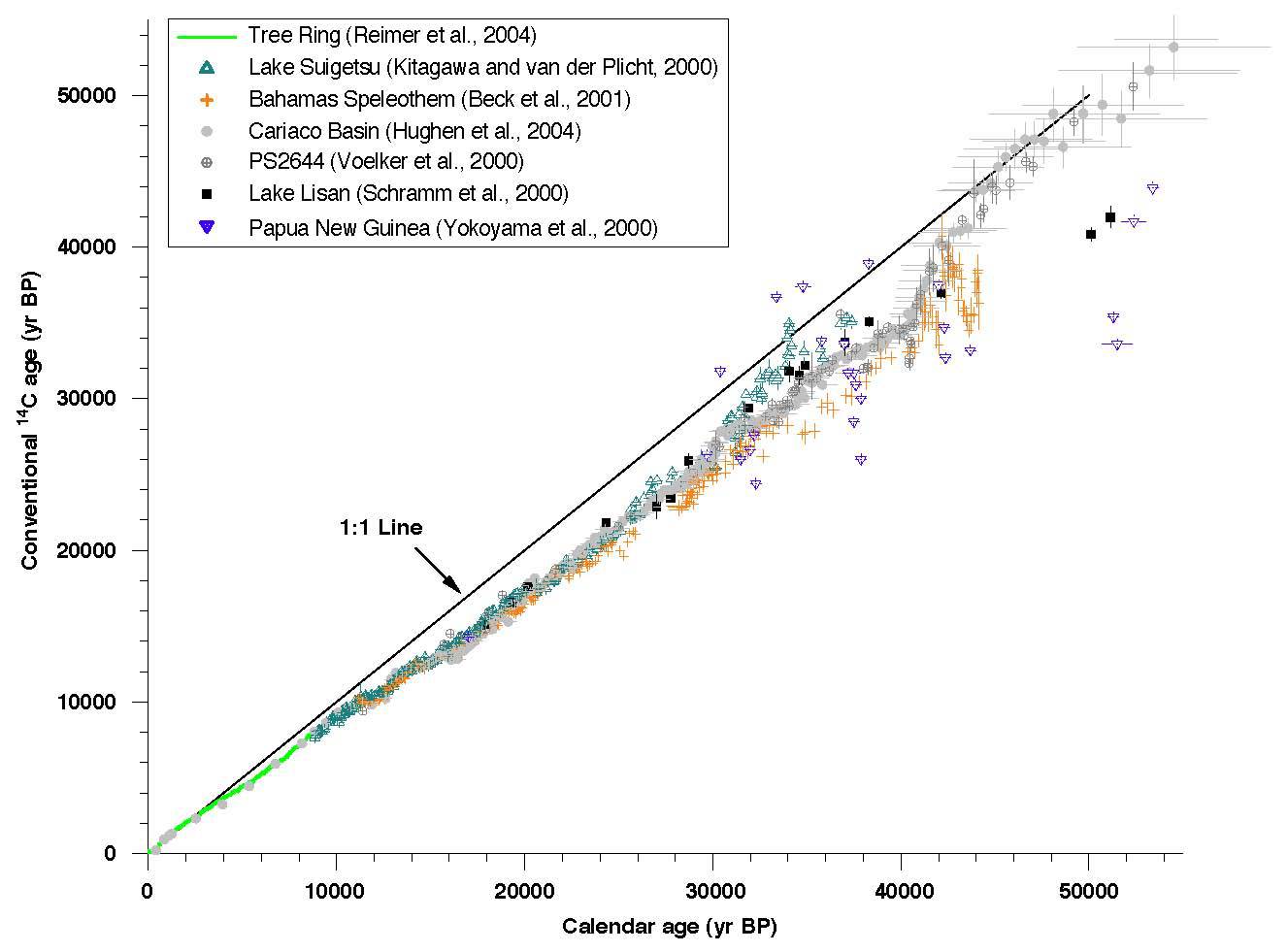
放射性炭素年代
紫色実線が較正前の放射性炭素年代と実年代の対応直線。
赤実線が較正後の放射性炭素年代と実年代の較正曲線。
http://upload.wikimedia.org/wikipedia/commons/b/b0/
Radiocarbon_dating_calibration.svg
だが実際は「前提条件」そのものが成立しない。宇宙線の影響や海洋に蓄積された炭素が放出されるなど、地球上の14C存在比率は常に変動することがわかった。そのために生じる測定誤差が、他の方法によるデータを使って補正されている(赤実線、較正曲線:後述)。
厳密には炭素14の生成量は地球磁場や太陽活動の変動の影響を受けるため、大気中の濃度は年毎に変化している。また、北半球と南半球では大気中の濃度が異なっている。
放射性炭素年代は、BP(Before PresentもしくはBefore Physics)で表記されるが、これは大気圏内核実験による放射線の影響をあまり受けていない1950年を起点として、何年前と実年代が表記される。
この測定原理を発見したウィラード・リビーは、炭素14の半減期を5568年として計算している。試料の分析結果から、2700から2400年前の約300年間は、新たな炭素14の追加が無かったことを意味するデータが得られているが、実際には生産量と同等な量の古い炭素が海洋から大気に放出されたと考えられている[要出典]。また、植物が炭素を固定する際に同位体選別と呼ばれる現象が生じ、植物の種毎に試料の炭素14濃度と年代には差異が発生する為、補正が必要である。同位体選別による炭素14の濃度差は、炭素13の濃度を精密に測定することで可能で、最大で400年程度の補正量となる。つまり、古い時代の生物の年代を正確に特定する為には、同じ種の現代の同位体選別量を正確に把握することも求められる。
大気中の炭素14量は、宇宙線の変動や、海洋に蓄積された炭素放出事件を反映して変動してきた。そのため、計測結果に誤差が生じている。後述の年縞堆積物および年輪年代により年代の較正が行われる。年輪年代では、およそ12600年程度までの放射性炭素年代値 (BP) と実際の年代の対応表が作られている[4]。年輪年代の及ばない古い年代は、およそ24,000年前までは、サンゴのU/Th(ウラン / トリウム)年代と照合されている。
較正曲線を用いて較正された年代値、つまり、炭素14年代を実際の年代に較正(基準に照らして正す)したという意味であり、西暦1950年を起点とした年数には calibrated(較正済み)を意味する「cal」をつけて「calBP」で表される。あるいは西暦紀元を基準とする場合は「calBC」ないし「calAD」と表す。較正年代は、暦年代 (Calendar year) とも呼ばれ、「実際の年代」という意味である。ちなみに、炭素14年代は「14C BP」となる。
AMS法

http://www.jaea.go.jp/04/aomori/ams/index.html
独立行政法人日本原子力研究開発機構(原子力機構)むつ事務所のAMS管理課では、平成9年4月にむつ事務所に導入されたタンデトロン加速器質量分析装置(JAEA–AMS–MUTSU)等の運転 • 維持管理及びAMS分析技術開発を行っております。
JAEA–AMS–MUTSUは、炭素及びヨウ素同位対比分析において世界トップクラスの性能を発揮しており、この技術基盤を活用し日本海及び青森沿岸海域を研究対象海域とした海洋における放射性物質等の移行挙動の解明に関する研究のほか、三内丸山遺跡からの出土試料や下北半島の埋没林の年代測定を行うなど、地域の歴史や自然史解明の一助も担ってまいりました。
現在は共用施設として、あらゆる分野における分析を行っておりますので、皆様のご利用をお待ちしております。
加速器質量分析装置(AMS)は、タンデム型加速器と質量分析計を組み合わせた分析装置です。B線計測などの従来の放射線計測法では、対象とする放射性核種自身の放射壊変により発生する放射線を計測するものでしたが、AMSでは、試料に含まれる同位体(炭素であれば12C • 13C • 14C)を物理的に分離し、その原子一つ一つを計測するため、試料の重量にして従来の約1000分の1以下の試料量で短時間で測定することができます。1試料あたりの測定は、炭素(14C)の場合は、30から60分程度の計測で従来法と同程度の精度で測定ができます。ヨウ素(129 I )の場合は、90から120分程度の計測で従来法では測定できない低レベルまで測定することができます。
平成9年4月にむつ事務所に導入されたAMSは、オランダの High Voltage Engineering Europa B • V 社製の小型タンデム加速器(3MV)を用いた新世代型の装置で、炭素及びヨウ素同位体比の測定が可能な2つのビームラインで構成されます。
本装置は、イオン源 • イオン入射システム部 • タンデム加速器部 • 高エネルギーイオン質量分析部から構成されています。
http://www.jaea.go.jp/04/aomori/ams/outline.html
イオン源
イオン源は、セシウムスパッター型イオン源です。測定試料はアルミニウム製の円柱状ホルダーの先端にプレスして詰め込まれ、ターゲットホルダー(カローセル)に59個まで装着することができ、コンピューターにより自動的に連続測定できます。
イオン源の原理は、高温に加熱されたタングステンフィラメントにセシウム(Cs)が接触しイオン化され、ターゲット電圧7KV、引き出し電圧35KVに印加された試料の中心に向かって加速照射されます。Csイオンにより試料をスパッタし、試料が表面から離脱する際に負にイオン化され35KeVのエネルギーを持ってイオン入射部に入射されます。また、コンピューターにより試料のCsイオンスパッターによるクレーター(穴あけ)効果を抑え測定精度の向上を図っています。
炭素の放射性同位体である14Cを測定する場合、同じ質量数を持つ窒素(14N)イオン、あるいは炭素の安定同位体(12C)と水素が結合したCH2イオン等が測定を妨害しますが、AMSでは最初に負のイオンをつくるときに電子親和性のない妨害元素(窒素)のイオン化を抑える等測定精度の向上を図っています。
イオン源
イオン入射部
イオン入射部では、イオン源で発生したイオンのうち、測定対象となる質量のイオンを分離して加速器に導入します。炭素用のイオン入射系は、高精度の測定を実現するためのリコンビネーターと呼ばれる入射系を採用しており、4台の45º 電磁石と2台の静電型スロットレンズから構成されます。リコンビネーターは炭素の3つの同位体(12C • 13C • 14C) を同時に入射させることにより、装置に起因する同位体効果を排除しています。初段の2台の電磁石により質量数12 • 13 • 14のイオンが約2㎝間隔に質量分離され、同時に不要のイオンが除去されます。3つの同位体が空間的に分離された軌道を通過するため、12Cのビーム強度を高速回転するチョッパーで約100分の1に低減し、加速器の負担を軽減し、発生するX線を抑えるとともに、14Cのビーム強度を相対的に上げ、分析時間の短縮を図っています。後段の2台の電磁石は、3つのビームを1つに収束し、タンデトロン加速器に入射します。
ヨウ素用のイオン入射系は、入射電磁石が内蔵されたバウンサー機構により、電磁石の磁界を変化させずに高速でイオンの加速エネルギーを変化させ、個々の質量を持ったイオンを逐次透過させ加速部に入射することができます。
イオン入射部
タンデム型加速器部
タンデム型イオン加速器は、高電圧発生部、Q–Snoutレンズ、低エネルギー加速管、荷電変換部、高エネルギー加速管、四重極型レンズにより構成されています。
高電圧発生部の昇圧方式は、信頼性の高い対称型コッククロフトウォルトン型高電圧発生方式を採用し、高電圧ターミナル部(荷電変換部)を最高3MV(300万ボルト)まで昇圧することができます。14C及び129 I 測定中は2.5MV(250万ボルト) に保持されます。
Q–Snoutレンズは、イオン入射部より導入されたイオンを荷電変換部に収束させるためのオートフォーカスレンズです。
加速管はチタン電極と絶縁物を接合したもので、高いコンダクタンスを得るために従来機種に比べ約40%ほど大きい直径(32㎝)の加速管が使用されています。この大口径の加速管により真空排気特性が向上し、高電圧ターミナル近傍で良い真空度が得られる設計となっています。低エネルギー加速管では、負イオンが荷電変換部に向けてターミナル部の電圧分加速されます。
荷電変換部に入射された負イオンは、荷電変換部内のガス(Ar)と衝突することにより電子を剥ぎ取られて正イオンに変換されます。荷電変換された正イオンは、高エネルギー加速管内でさらに加速され、ターミナル部電圧に荷電数を掛けた値のエネルギーを持ち加速管から放出され四重極レンズで高エネルギー質量分析部に向けて収束されています。
タンデム型加速器部
高エネルギーイオン質量分析部
高エネルギー質量分析部では、通常の質量分析装置と同様に加速されたイオンを磁場及び静電場によって質量分離します。
炭素用の高エネルギー質量分析部は、110º 電磁石、90º 電磁石、静電アナライザー、重イオン検出器から構成されています。加速器を通過した12C、13C、14Cイオンは、初段の110º 電磁石により質量分離され12C、13Cイオンビームはファラディーカップにより電流測定されます。14Cイオンは静電アナライザー、90º 電磁石で分析され、重イオン検出器でパルス測定されます。
ヨウ素用の高エネルギー質量分析部は、115º 電磁石、静電アナライザー、飛行時間(TOF:Time of flight)型検出器から構成されています。加速部を通過したヨウ素イオンは、115º 電磁石により質量分離され、127 I イオンビームはファラディーカップにより電流測定され、129 I イオンはTOF型検出器で測定されます。
高エネルギーイオン分析部
http://www.jaea.go.jp/04/aomori/ams/details.html
http://www.radiocarbon.org/IntCal04%20files/Intcal04wcaption.pdf
1976年 加速器質量分析法(AMS)を用いた14C年代測定法の開発
1970年代末に開発された分析手法。加速器で炭素14を直接数える方法 AMS(Accelerator Mass Spectrometry = 加速器質量分析計)で、必要な試料量(1mg程度)、測定時間(30分 - 1時間程度)共に大幅に改善され、ベータ線計測法と比較し高精度化・高効率化された。また約6万年前まで測定可能となった。装置の小型化に伴い多くの施設で入手可能なレベルになっている。
加速器質量分析 (Accelerator Mass Spectrometry, AMS)
加速器を利用し、物質を通過する際のエネルギー損失率の差などを利用して同重体などを除去し、特定の原子のみを計測するものである。考古学での炭素年代測定などに利用される。加速器を利用するため、非常に大掛かりな装置となる。
遺跡などから発掘されたものの中から有機物を含むものを試料とします。
例えば、甕や土器の内包植物、吹きこぼれ、おこげ、また、骨などからコラーゲン、衣服、から試料を採取します。これらを化学処理によって、現代炭素の汚染を除去し、酸化・二酸化炭素・ 還元・グラファイトというプロセスを経て試料中の炭素としてAMSで測定します。このグラファイトは1mg程度で充分な量なので試料自身の量も少なくてすみます。
現代炭素1g中には約600億個の14Cが存在します。20,000年前の試料でも約50億個以上が存在します。従来のβ崩壊を測定する方法では現代炭素でも統計誤差1%を得るには約12時間の連続測定が必要です。AMSでは14Cを直接計るので非常に短時間で測定できます。
炭素中には12C、13C、14Cがそれぞれ99%、1%、10-12 ~ 10-14存在します。
イオン源に前述の試料から精製したグラファイトを充填し炭素の負イオンを発生します。これを磁場を使った分析装置(電磁石)と電磁石中のチャンバーに電圧をかけ、12C、13C、14Cを逐次加速器に入射します。例えば、12Cを300μsec、13Cを900μsec、14Cを100msecと言うサイクルを繰り返し積算します。加速された炭素イオンはイオン源でCH、CH2等の同重体があっても加速ターミナル内の荷電変換(通常はアルゴンガス)で分解され炭素イオンとなる。これを再度、電磁石と静電アナライザーで分析し検出器でそのエネルギーと個数を計数します。検出器には、測定すべきイオンの他に、他のイオンやイオン源からの分子イオンが散乱されてたまたま検出器に到達してしまうものがあるがこれらはエネルギーの違いで分離することができます。
測定試料と標準サンプルと同時に測定され、これらの比較により14C/12C比、13C/12C等から規格化され、14C AMS年代が決定されます。
加速器質量分析 法 (AMS = Accelerator Mass Spectrometry) では、分析したい試料自身を加速してイオンビームとし、 質量分析を行う。この方法は特に、極微量の長半減期放射性同位体の検出に威力を発揮する。
放射性同位体の定量には、その崩壊に伴って生成する放射線 (ガンマ線やベータ線) を測定する方法が 存在する。これは、放射線の生成率が、放射性同位体の存在量に比例するという原理を用いて試料中の 放射性同位体の量を知る方法である。しかし同時に、放射線生成率は、核種の半減期に反比例するので、 半減期が何千年、何万年という長半減期の核種の測定の効率は悪くなる。
一方、AMS では、崩壊しないで残っている方の原子核の個数を直接測定対象とする。したがって、半 減期の長い核種ほど、放射線計測よりも、AMS の方が有利となってくる。例えば、半減期 5,730 年の 14C を測定する場合、感覚としては、β カウンティングのために必要なサンプル重量がグラムオーダー であるのに対し、AMS ではミリ・グラムオーダーで十分である。これにより、それまで手の出せなかっ た、貴重な考古・人類学試料の 14C 年代の測定が次々と行なわれるようになった。
まず測定したいサンプルそのものを適当な化学種としてイオン源に装填し、1 価の負イオンビームと して引き出す。イオン源としては、多くの場合セシウムスパッター型の固体イオン源が用いられる。し たがって、サンプルは固体粉末とされ、カソード形状に合わせてプレスされる。引き出されたイオンは、 入射電磁石により特定の質量のイオンのみを選んで加速器に入射する。タンデム加速器の場合、ターミ ナル部で荷電変換され、多価イオンビームとなる。加速器より出てくる高エネルギービームは、分析電 磁石により、特定の質量、電荷のもののみ選別される。最終的には、固体検出器などを用いて、高エネ ルギー粒子として検出する。
しかし、イオン源に装填した原子核すべてを検出できるわけではない。イオン源から最終検出器にい たる各過程で、さまざまな割合でロスがあり、最終的な検出効率は、10-2 ∼ 10-5 程度となる。この検 出効率は、核種や実験条件によって異なるため、正確に求めることは困難である。また、ある一回の測 定を行った際、イオン源に装填したサンプルのうちの何パーセントを消費したかを見積もることは極め て難しい。これらのことにより、着目した核種のみを検出しても、もともとのサンプルに含まれていた 量を推定することは困難である。
そこで、AMS では、測定したい同位体 (rare isotope) と同時に、安定同位体 (stable isotope) を定量 する。通常、安定同位体は測定したい同位体よりはるかに多く存在するため、電流として測定される。 両者は、サンプル中で、同一の化学種として存在するため、イオン化効率はほぼ等しいと考えられる。 したがって、どのような条件でも、加速されるイオン中の同位体比はもともとのサンプル中の同位体比 を反映していると考えられる。
AMS で得られる結果は、rare isotope の絶対量ではなく、stable isotope との同位体比である。
一般の質量分析法では、静電フィルター (エネルギー分析)、磁場フィルター (運動量分析) を利用して イオンビームの比電荷を分析する。これは AMS でも同じである。しかし、これらのフィルターだけで は、目的核種と同重の分子イオン、あるいは同重体核種を分離することは困難である。
たとえば、10Be を検出したい場合は、同重体 10B が、直接妨害核種となる。14C の場合は、14N が負 イオンを形成しないことから、イオン源としてセシウムスパッターイオン源等、負イオン源を用いる場 合には、同重体の妨害はほとんどない。しかし、13 CH- 、12 CH-2 などが同重分子として妨害となる。
また、イオン源で加速される粒子ビームは、どんな場合でもエネルギーが完全に均一ではなく、分布 を持っている。その場合、質量数の 1 つだけ異なる原子や分子がフィルターを通り抜けて最終検出器に 到達する可能性がある。例えば、目的核種より質量数が 1 だけ大きい核種 (あるいは分子) を考えてみる と、イオン源から引き出される際、そのエネルギー分布が低エネルギー側に裾を引いている場合、目的 核種と運動量が等しくなる成分が含まれる。この場合、入射電磁石のフィルターを通過してしまう。
これらに対して、加速器を用いて核子あたり MeV オーダーのエネルギーを与えると、薄いガスや薄 い膜を通過させることによって、容易に分子イオンを壊すことができる。分子イオンは壊れてしまえば 質量が異なるので、その後のフィルターで容易に分離できる。加速器としてタンデム加速器を用いる場 合には、荷電変換装置として、ターミナル部 (最も電圧の高くなる部分) に薄いガスを流すキャナル (ス トリッパーキャナル) や、炭素膜を備えており、この部分で荷電変換と同時に分子イオンが破壊される。
また、核子あたり MeV オーダーのエネルギーのイオンビームに対しては、最終検出器でエネルギー を測定することが可能である。たとえ目的核以外の原子が最終検出器に到達したとしても、エネルギー の違いで、容易に分離できる。さらに、物質中でのエネルギー損失が原子番号 Z によって異なることを 利用すれば、最終検出器で、媒質中のエネルギー損失と残留エネルギーの両方を測定することによって 同重体を分離することができる。
AMS の出現によって、それまで測定できなかったサンプルのデータが得られた。測定結果が、それ までの定説を覆すような場合も少なくない。そのような場合、AMS 測定の信頼性そのものに対する疑 問の声が上がることが少なくない。AMS 測定においては、イオン源におけるイオン化や、加速器ター ミナルにおける荷電変換など、確率的な素過程に支配されているため、得られた計数に伴う統計誤差が 存在する。また、加速器の透過率や荷電分布の同位体間での違いから系統的な誤差を生じる可能性もあ る。微量なサンプルを扱うことが多いため、サンプル作製過程において、コンタミネーションや同位体 効果の起きる可能性もある。これらを詳細に吟味し、得られるデータの信頼性を維持すること、これが AMS 研究施設をあずかる科学者の仕事であると考えられる。
松﨑 浩之(東京大学原子力研究総合センター タンデム加速器研究部門)
AMS(加速器質量分析) とは
http://malt.n.t.u-tokyo.ac.jp/PastInfo/10thSympo/matsuzaki.pdf
樹木年輪年代法

The growth rings of a tree at Bristol Zoo, England.
http://upload.wikimedia.org/wikipedia/commons/7/7d/Tree.ring.arp.jpg
標準パターン(下)に試料のパターン(上)が一致する
http://www2.odn.ne.jp/hideorospages/yamatai03.html
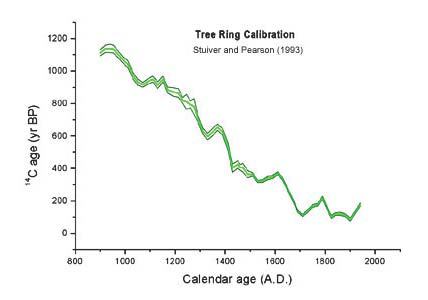
http://www.physics.arizona.edu/ams/education/calibrate2_1.htm
Radiocarbon ages of wood from tree rings of known age have been replicated from a number of laboratories around the world with high precision.
Δ(Age) ≈ 0.2% or ±16 years
Plots of true age (from tree rings) vs. radiocarbon age allow one to calibrate radiocarbon ages.
年輪年代法 モノサシは年輪
年輸年代法(正しくは年輪年代測定法)は、1920年代にアメリカの天文学者、A・E・ダグラスによって創始され、欧米ではすでに建築史や美術史など、さまさまな分野で実用化されています。
その原理は、いたって簡単。樹木の年輪が毎年一層ずつ形成されることを利用しています。
樹木の年輪というのは気象条件に左右され、生育のよい年と悪い年、つまり年輪の幅が広い年と、狭い年があります。
その変化を何十年という期間で追っていくと、年輪幅の変化がパターンとなって現れてきます。木材の種類によって、共通するパターンが見えてくるのです。 そのパターンを過去へ過去へと延ばしていくと、古代までひとつながりの年輪のパターンができあがります。いってみれば、樹木の年輪によって、過去何百年、何干年というモノサシができるわけです。
このようにして作成された長期の年輪パターンと、遺跡などから出土した木材の年輪パターンを照合することで、古代の木材が切り出された年が1年単位で判明する。そういう画期的な年代測定法です。
しかし、年輪年代法で測定するためには、出土した古代の木材の一番外側の年輪(最外年輪)まで残っていることが基本的に必要です。
年輪年代法は日本にも適用できる
年輪年代法はしかし、日本では長い間使いものにならないといわれてきました。
ヨーロッパやアメリカの乾燥地帯のように気象条件の厳しいところでは有効でも、日本のように気候が温暖なうえに、地形も複雑、おまけに地域ごとに微妙に気候が変化するようなところでは、ほとんど役に立たないと考えられたからです。
池上曽根遺跡出土「柱根12」
日本でもこれまで、大正時代以来何度か実用化が模索されてきましたが、十分な成果を得るまでに至りませんでした。
よく知られたところでは、戦後間もないころ、奈良の法隆寺の五重塔が解体修理されたとき、年輪年代法によって法隆寺の建設年代を知ろうという試みがありましたが、このときは残念ながら、失敗しています。
ところが、1980年代から新たな動きが始まります。奈良国立文化財研究所(以下、奈文研)の光谷拓実さん(当時32歳)が、年輪年代法の本格的な研究を開始します。
光谷さん自身は、もともと植物の専門家で、考古学者でも歴史家でもありません。そういう部外者ともいえる人が、日本の古代史にとってきわめて重要な役割をはたすのです。
年輪のデータを徹底的に取る
光谷さんがまず最初にしなければいけなかったのは、徹底的に樹木の年輪のデータを取りつづけることでした。各地の営林署や営林局へ行って、木材の試料を集めてきます。
まず、現生木のデータを徹底的に調べます。ひとつの種類の木だけでも複数の試料が必要です。それを何種類もの木に広げていきます。
年輪を読み取る光谷拓実さん 日本では本当にダメなのか、それを知るためにもデータを取りつづける以外にありません。年輪の測定には10ミクロン、つまり100分の1ミリというような単位が必要です。
そのくらい細かなデータでなければ、年輪幅の変化を追っていけないからです。それをグラフにしていきます。くる日もくる日も、そんな作業の繰り返しでした。
そして3年が過ぎたころ、ようやく最初の成果があらわれてきました。日本のような複雑な気象条件では、年輪年代法は不可能と長年いわれてきましたが、日本の樹木でも共通するパターンが見えてきたのです。
古くから建築材として使われてきたヒノキやスギを調べると、たしかに年輪の幅は個々の木で違っていますが、何年分という期間で照合すると、地域に関係なくパターンが共通することがわかってきました。
しかも、アメリカでは15年分くらい、ヨーロッパでは30年分ほどでパターンの照合ができますが、気候の違う日本では100年から200年分でやっと正確な照合ができることがわかりました。
これは大きな前進です。日本でも年輪年代法が使える可能性が出てきたわけです。
日本でこれまで年輪年代法が成功しなかったのは、これほど徹底したデータの収集が行われなかったからでした。
弥生時代までモノサシが届いた !
研究の次の段階は、年輪のパターンを過去へ過去へと、どんどん延ばしていく作業です。これには古い寺院の修理部材や、中世、近世の遺跡から出土する試料が役にたちました。
驚いたことに、それらの年輪を調べても、パターンはピタッ、ピタツと一致します。
同時に、奈文研には平城宮の発掘で出土した大量の古代の木材が保管されており、そこからもデータを集めました。
こうして研究を始めて6年が過ぎたころ、ようやくはっきりとした成果が出てきました。過去2000年分、つまり現代から弥生時代まで届く年輪のモノサシが、やっとできあがったのです。
日本では不可能といわれていた年輪年代法に、ついに実用化のメドがたったのです。
標準パターン(下)に試料のパターン(上)が一致する
年輪年代法は、1985年11月、紫香楽宮(しがらきのみや〕跡を特定するという画期的なデビューをします。
滋賀県の宮町遺跡から出土した柱根の年代を測定すると、『続日本紀』の紫香楽宮の記述とドンピシャリ、しかも建設された季節まで一致したのです。
これによって、紫香楽宮跡は宮町遺跡であることが科学的に明らかになりました。その後、さまざまな分野に活用されはじめ、東大寺の仁王像や、法隆寺の五重塔の心柱の年代測定、さらに円空仏の真贋論争にまで活用されました。
ところで、最も期待されていた弥生時代の年代を決めるというテーマは、長い間、足踏み状態が続いていました。年輪のパターンはすでにスギやヒノキで紀元前1000年ごろまで延びていましたが、試料となる弥生時代の木材が出土しなかったからです。
画期的な弥生時代像に色を失う考古学者
いよいよ弥生時代の遺跡から木材が出土するのは、研究を始めて十数年が過ぎた1995年の秋です。
滋賀県の二の畦・横枕(にのあぜ・よこまくら)遺跡というところから弥生中期の井戸が発見され、そこに残っていた井戸材の年代を測定しました。
結果は思いがけないものでした。年輪年代法で得られた年代は、考古学者が考えていた年代よりもなんと、100年以上古かったのです。
紀元後1世紀ごろの遺跡と見られていたのが、年輪年代法では紀元前1世紀の前半と出ました。
当時、定説とされていた年代観とは、大きく違います。
しかし、不幸なことに、考古学者の間には、年輪年代法は日本では無理という考えが、まだまだ根強く残っていました。そのため、当時は年輪年代法で得られた結果に不信感を抱く研究者も少なくなかった、といわれています。
池上曽根遺跡の大型建物と井戸跡 けれど、翌年の池上曽根遺跡のケースでは、考古学者は色を失いました。池上曽根遺跡は、近畿を代表する弥生時代の遺跡で、過去に多くの有力な学者が発掘に参加してきました。
ちょうど前年には、この遺跡から大型建物跡が発見され、多数の柱根が残っていたことで注目を集めていました。
弥生の定点を定める
この大型建物は、有名な吉野ケ里遺跡の建物よりも200年ほど古い時代のもので、中国の影響を思わせるように、建物の向きは東西南北にきっちりと合わせて建てられています。
しかも、出土した柱根のなかには最外年輪まで残っているものがありました。
さっそく光谷さんが柱根の年代を測定したところ、やはり思わぬ結果が出ました。
紀元後1世紀後半と予想されていた建物の年代が、年輪年代法では紀元前52年と出ました。定説よりもやはり100年も古いのです。前年の二の畦・横枕遺跡のケースとまったく同じです。
考古学関係者はここで、年代論にまで踏み込んだ徹底的な検証を開始しました。じつは、年輪年代法の結果は、かねて少数の若手研究者が主張していた年代観にかなり近いものでした。「近畿の年代はもっと古いはずだ」という主張です。権威ある「定説」の前では「異分子」の扱いでしたが、もはや、彼らの主張を無視することも、年輪年代法の結果を無視することもできませんでした。定説を根本から再検討する必要があったのです。そして、出された結論は、まさに衝撃的でした。
これまでの定説を覆し、近畿の年代観を従来よりも100年も古くとり、先進の北部九州と並行する時間軸で捉えたのです。弥生時代の近畿と九州が、同じ年表に並んだ瞬間でした。
都出比呂志さんの案によるふたつの編年表。
1983年(左)のものと、1998年(右)のもの。
―拙著『卑弥呼の謎 年輪の証言』より―
年輪年代法によって、従来の年代観がガラリと変わったのです。
日本では使いものにならないとまでいわれた年輪年代法は、ここにおいて名実ともに認められることになりました。
年輪年代法が、これまで不確定だった弥生の年代決定にたいして、動かない定点を与えるという重要な役割を果たしたのです。
http://www2.odn.ne.jp/hideorospages/yamatai03.html
ウラン-トリウム年代測定法
ウランはアルファ崩壊、ベータ崩壊を繰り返して、安定な鉛の同位体になる。この間、トリウム、ラジウム、ラドンなど科学的な性質の異なる核種を経過する。崩壊系列の途中の核種の半減期は一般に短いが、
234Uの半減期は24.5万年、230Thの半減期は7.5万年と比較的長い。
しかも、酸化ウランが水に溶けるのに対して、トリウムは水に溶けず、すぐに沈澱する。
この2つの特徴を利用したのがイオニウム(Io: 230Thのこと)年代測定法である。
河川は陸上の岩石を浸食し、その成分を海に運んで来る。この中に、ウランとトリウムも含まれている。
ウラニルイオン(UO2)2+は海水に溶け、海水中に長時間滞留する。平均滞留時間は約50万年である。
トリウムは海水に溶けず、約300年の平均滞留時間で海底に沈澱する。従って、海底堆積物表層部にはトリウムが多く含まれることになる。
陸上の岩石に含まれるウランは放射系列の途中の核種で半減期がもっとも長い234Uでも半減期は25万年を切るのであるから、地質年代にくらべればほぼ一瞬のうちに放射平衡に達している。つまり、238Uの崩壊で単位時間に生じる234Uの原子数と234Uの崩壊で単位時間に生じる230Thの原子数は等しく、また230Thの崩壊で単位時間に生じる226Raの原子数も等しい。大本の供給源である238Uの半減期は45億年と桁違いに長いので、短期間では238Uから単位時間に供給される原子の数は変化しない。230Thが放射崩壊で減る分は234Uの崩壊によって補われ、234Uが崩壊によって減少する分は、238Uの崩壊によって補われる。
ところが、海底堆積物に関しては、上に述べたように、230Thの供給源である234Uおよび大本の供給源である238Uが海水中に長期間滞在するため、海底堆積物中にお沈澱した230Thは供給源を失うことになる。
測定に際しては、230Thと232Thの放射能の比を測る。この2つは同位体なので、化学的には同じ行動をする。海水中におけるこれら2つの同位体の比が変わらなければ、海底に沈殿した時点での比も同じである。時間が経つに連れて230Thの放射能は減少するが、232Thの放射能は減少しない(半減期140億年!)。比で測定する利点は、同位体比の方が230Thの絶対量より変動の影響を受ける程度が少ないためである。(何かの原因で230Thの沈殿量が5%減ったとしても、232Thの沈殿量も5%減ると期待できる。)
説明を簡単にするために、堆積物にUは全く含まれないとしよう。この場合、堆積物中の230Thは半減期7.5万年で減少して行くことになる。海底堆積物の表面にはいつの時代でも同じ濃度の230Thが含まれるとしよう。もし、1メートルの深さの堆積物中の230Thの濃度が50%になっているならば、7.5万年前に堆積した堆積物と考えられる。このようにして、堆積速度を見積もることが可能になる。
http://ksgeo.kj.yamagata-u.ac.jp/~kazsan/class/chronology/ionium.html
Multicollector - inductively coupled plasma - mass spectrometry (MC-ICP-MS) provides a unique opportunity to perform precise isotope ratio measurements for a wide range of elements.
Multicollector - magnetic sector double focussing - inductively coupled plasma - mass spectrometry (MC-MS-ICP-MS).
放射性炭素(14C)年代測定法の上限4~5万年やウラン・トリウム(230Th0/234U)法の上限35万年
福井県水月湖(すいげつこ)の湖底地層の年縞(ねんこう)
福井県に三方五湖(みかたごこ)と呼ばれる五つの湖がある。その中で最大面積を有する水月湖(すいげつこ)は、近年とみに、環境考古学、古気象学、考古学的遺物の年代決定に重要な湖として世界的な注目を浴びている。
まず水月湖は二重底の湖として知られる。水深0~6mの湖水上部は淡水であり、水深7~40mの湖水下部は汽水である。汽水とは海水と淡水が混在した状態のことだが、水月湖の湖水下部にあるのは、硫化水素を含むが酸素を含まない死の水である。
水月湖は水深が深く、湖内に直接流れ込む大きな川がない。水路工事が行なわれ、南の三方湖(みかたこ)や北東の久々子湖(くぐしこ)とつながっている。三方湖からは淡水が、久々子湖からは汽水が流れ込むようになった。比重の重い汽水は湖底に滞留している。

http://upload.wikimedia.org/wikipedia/commons/1/18/
Mikata_five_lakes_Aerial_photograph.1975.jpg より。
湖の表面に強い風が吹くと表層の淡水は撹拌されるが、湖底の汽水はかき乱されることがない。その結果、下の汽水は空気に触れることなく、酸素を含む上の淡水と混じり合うことがない。湖底の汽水は、有機物分解時に酸素が消費しつくされ、硫化水素を多く含む無酸素状態となった。
湖底には、春から夏はプランクトンや珪藻の死骸が堆積して白色の層ができる。秋から冬は粘土鉱物が堆積して黒色の層が重なる。この白と黒のバーコードタイプの縞模様が年縞(ねんこう)と呼ばれ、白黒一組が一年に相当する。
湖底の汽水に流れがないため堆積物はかき乱されることがない。酸素のない湖底に生物が生息しないため堆積物が食い荒らされることもない。こうして、水月湖の年縞は一枚ずつきれいに積み重なった状態で保たれていた。
湖周辺の断層の影響で、湖底が沈降していっており、堆積物が積もっても湖が埋まらないという特別な状況が重なっていた。それゆえ、水月湖の年縞は「奇跡の堆積物」と呼ばれるようになった。
水月湖のボーリング調査が実施され、過去十六万年分の連続した年縞を採取できた。二〇〇六年に採取されたサンプルということで、SG06と呼ばれた。
年縞SG06を元に精度の高い環境変動のデータを得ることができた。そればかりでなく、堆積物の14C/12Cの比率をAMS法で調べることで、約五万年分の放射性炭素年代測定を行なって較正曲線が描くことができた。
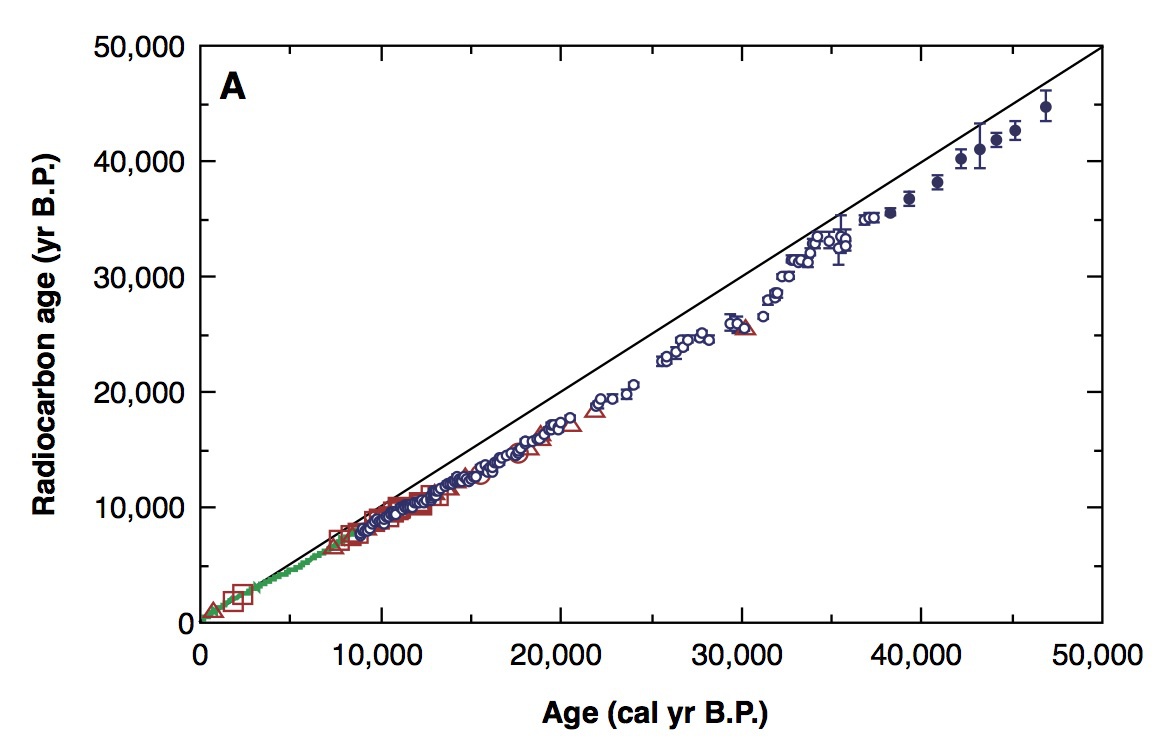
Kitagawa, H. & van der Plicht, J., Science 297 (1989) 1187-1190、Fig.1
この図は、水月湖の年縞から得られたサンプルを放射性炭素年代測定し、他の方法すなわち年輪年代法や海洋生物の珊瑚を用いた年代決定法によく一致することを示している。
本図における他のサンプルとは、年輪年代法(緑色実線)ならびに珊瑚サンプルであり、使われた珊瑚はそれぞれパプアニューギニア(□)、ムルロア環礁(○)、バルバドス(△)産である。
水月湖の年縞によるデータはエラーバー付きの小さな紺色○印である。
三万八千年より前はデータが大きく変動していた。そのため、一定速度で堆積した場合を推定した値が表示されている(エラーバー付きの小さな紺色●印)。
データの大きなバラツキは、氷河期だった当時、大気中の14C含量が著しく変動していたことを示唆している。地球規模の環境変化や宇宙線による同位体生成率の変化に関係しているかもしれない。
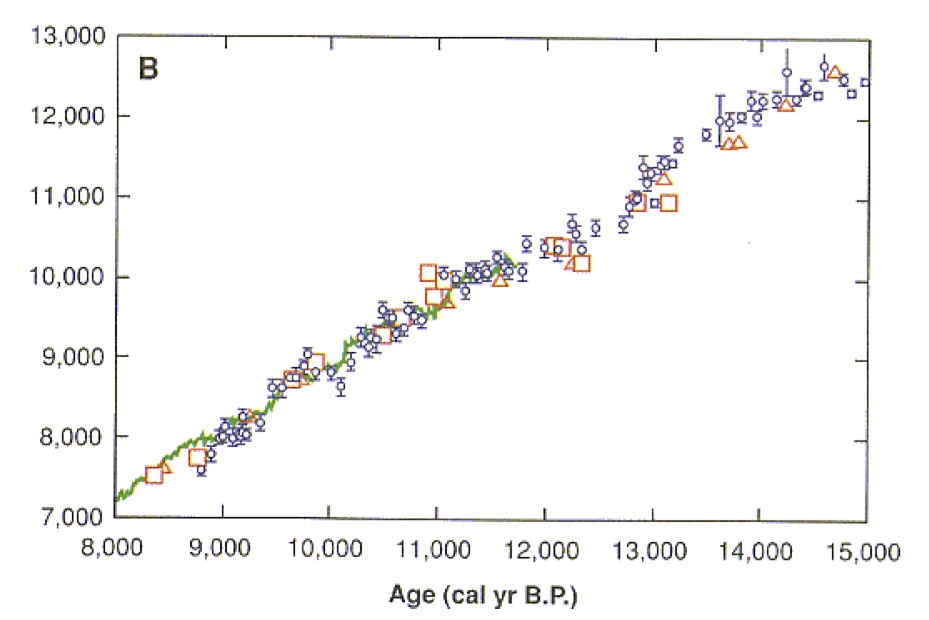
Kitagawa, H. & van der Plicht, J., Science 297 (1989) 1187-1190、Fig.2
この図も、水月湖の年縞から得られたサンプルを放射性炭素年代測定し、他の方法すなわち年輪年代法や海洋生物の珊瑚を用いた年代決定法によく一致することを示している。
本図における他のサンプルとは、年輪年代法(緑色実線)ならびに珊瑚サンプルであり、珊瑚はそれぞれパプアニューギニア(□)、バルバドス(△)産のものが使われたデータである。
水月湖の年縞によるデータはエラーバー付きの小さな紺色○印である。
Fairbanks, R.G., et al., Quaternary Science Reviews 24 (2005) 1781–1796、Fig.1
この図も、年縞による年代決定法(▲緑色三角形)が、他のサンプルを使った年代測定法によく一致していることを示している。
本図における他のサンプルとは、年輪年代法(緑色実線)、バハマ諸島の洞穴内生成物(+オレンジ色十字)、パプアニューギニア珊瑚(▽紫色逆三角形)、リサン湖湖底堆積物(■黒四角)、ヴェネズエラ沖カリアコ海盆(●薄灰色丸印)の浮遊性有孔虫の海底堆積層、アイスランド北方沖の海底(濃灰色丸十字)の浮遊性有孔虫の海底堆積層である。
これらの結果、誤差は約五万年で一七〇年程度と非常に高精度であることがわかり、水月湖年縞からのデータは、地質学的年代決定の事実上の世界標準として認められた。
穏やかな水底(淡水および海水)に堆積した物質の縞模様のうち、年毎に一定の層序を保ち堆積した物は年縞堆積物と呼ばれ正確な年代を決定できる。主な、採集地は以下である。
ドイツ中部のアイフェル地方にある湖沼群(完新世と晩氷期)
ベネズエラ沖のカリアコ海盆の堆積物(晩氷期以降の時代)
福井県水月湖約7万年分[4]
較正曲線を使って、どのように実測14C年代から較正歴年代を推定するかを、模式的に表したのが次の図である。
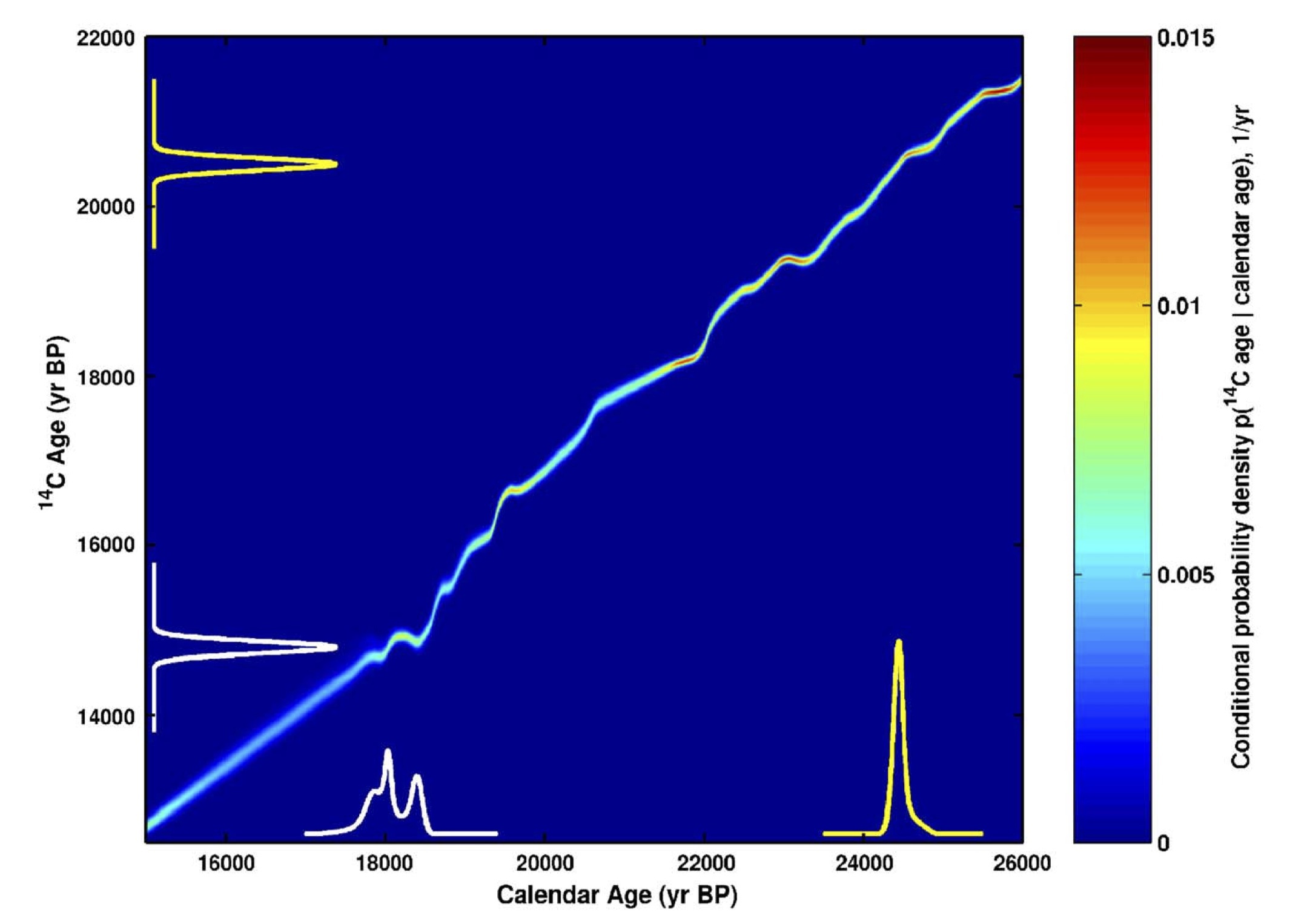
Fairbanks, R.G., et al., Quaternary Science Reviews 24 (2005) 1781–1796、Fig.4
図の縦軸が実測14C年代、横軸が較正歴年代、図に描かれている曲線が較正曲線である。例えば縦軸の二万五百年程度のところに実測14C年のスペクトラムが描かれているが、その一点一点を横軸方向にのばし、較正曲線と交差した点から縦軸方向に下に向かってのばす。
その直線がX軸と交わったところに描かれているが、較正歴年代のスペクトラムである。その結果、較正歴年代は約二万四千五百年と計算されることになる。
こうしてまとめたように、高精度質量分析計が導入され、水月湖の年縞など堆積物のデータを用いることにより、考古学的遺物の正確な年代測定が可能となった。
すなわち土器に練り込まれている土の解析により土器が作製された年代が、焦げ付いた食物の解析により土器が使用されていた年代が、プラスマイナス数十年のオーダーで推定できるようになった。
単なる思い込みや推論ではない。ワンランクもツーランクも上のレベルで、論理の展開が可能になった。より真実に迫れるようになったのである。
日本人のルーツ(Ⅲ)世界最古16500年前の土器片
2013.10
世界最古16500年前の土器片 
大平山本I遺跡(おおだいやまもといちいせき)出土の土器片
世界最古16500年前の土器片

世界最古の土器が日本で発見される
日本で最初に測定されたのは、1950年・1955年に調査された夏島貝塚の縄文時代早期の層から出土したカキ殻と木炭であった。1959年3月と6月に、ミシガン大学から杉原荘介に、炭素14年年代法による年代値は、貝殻BP9450±400と木炭BP9240±500であったことが報告された。この測定の結果、縄文時代早期に9500年前という年代がはじめて示され、縄文土器が世界最古の土器文化である可能性が指摘される一方、日本の考古学者の多くを驚愕させた。また、測定を依頼した芹沢長介らと、大陸で出土した遺物の年代から3000年前と主張する山内清男との間で論争が起きている。
青森県東津軽郡外ヶ浜町の大平山元I遺跡の縄文時代草創期の土器製作時期が、通説より4500年も古い(早い)1万6500年前と1999年4月に発表された。この実年代は、ワシントン大学のスタイヴァーらが炭素14年代を年輪年代や珊瑚年代を使って暦年に換算する国際較正曲線 (INTCAL 98) を使ったものである。また、弥生時代の開始期は通説では紀元前5 - 紀元前4世紀ごろであったが、2003年3月の国立歴史民俗博物館の発表では約500年古い(早い)約3000年前(紀元前10世紀終頃、つまり、九州北部の弥生時代早期が前949年 - 915年から、前期が前810年頃から、中期が前350年頃から、それぞれ始まった。)に遡る結果が出た。2003年5月の日本考古学協会総会での報告は、衝撃、当惑、賛成、反発などとともに拒否、嘲笑などに覆われた。しかし、その後も試料収集と測定は進められ、その成果が期待されている。
日本各地の交流、交易
日本列島と朝鮮半島の交流、交易
縄文人の豊かな生活様式(1)
日本人のルーツ(Ⅳ)水田稲作は紀元前10世紀頃から
2013.10
水田稲作は紀元前10世紀頃から 
水田稲作跡が発見された福岡県板付遺跡の航空写真
水田稲作は紀元前10世紀頃から

二千三百年前、稲作開始説
プラントオパール
縄文土器の胎土に発見される稲のプラントオパール
三千年前、水田稲作開始
六千年前陸稲栽培開始
稲DNAのSSR多型から日本の稲作がどこから到来したか判明
縄文人の豊かな生活様式(2)
日本人のルーツ(Ⅴ)グレートジャーニー
2013.10
グレートジャーニー 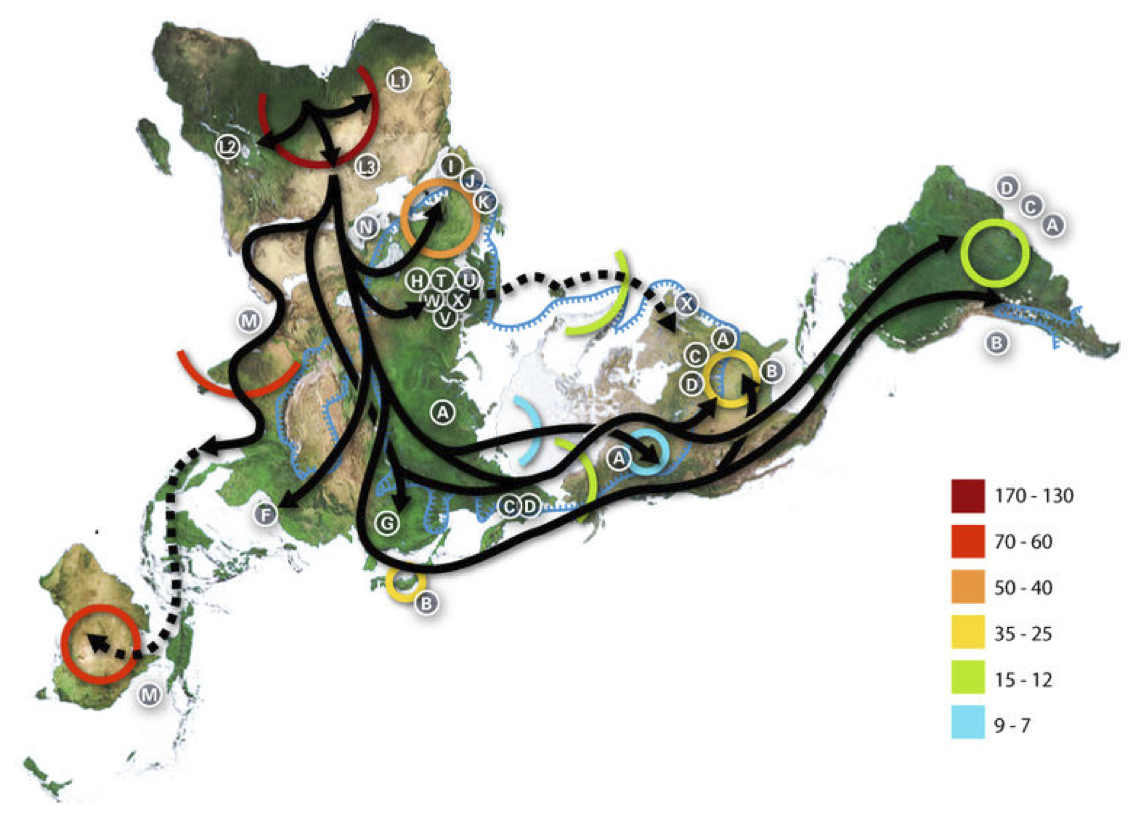
ミトコンドリアDNA解析によるイヴの子孫たちの移動と分布
Wikipediaによる(CC BY-SA 3.0)
http://creativecommons.org/licenses/by-sa/3.0/deed.en
ダイマクション地図についてはこちらを参照
本稿の文献はこちらを参照されたい。
グレートジャーニー
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
このページのまとめ
1)細胞の中の小器官ミトコンドリアにもDNAがある。
2)母から娘、またその娘へと、代々薄まることなく受け継がれる。
3)突然変異が起きるとそれも受け継がれる。
4)代々積み重なった変異の多様性はグループ分けされ、ハプログループと呼ばれた。
5)ハプログループの研究から、分子人類学が勃興した。
6)新人の単一起源説、アフリカ起源説が提唱され、他地域起源説は全面否定された。
7)イブの子孫が出アフリカを果たし、世界各地に移住して行った。
8)その移動はグレートジャーニーと呼ばれる。
9)ミトコンドリアDNAハプログループは、L、M、N、Rの各クラスターに別れる。
10)日本人のハプログループ構成頻度は、Mが61%、Nが14%、Rが19%である。
11)大陸北部と日本には共通した亜型が存在する。
12)大陸南部には日本人に多い亜型とまれな亜型が並存する。
13)それぞれ北方系、南方系と呼ばれ、日本列島に渡って来たと考えられた。
14)Y染色体DNAハプログループの研究が始まるまで、その考えが主流だった。
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
Ⅰ)ミトコンドリアDNAハプログループの登場と新人の単一起源説
出土した頭蓋骨など人骨を計測して特徴を解析し、これが○○人、あれが□◇人と分類していたのが形態人類学である。形態人類学から導き出された世界観を、根底から揺さぶる研究領域が勃興した。分子人類学である。
それまで「多地域起源説」が主流だった。一〇〇万年以上前にアフリカを旅立った原人が各地で独自の変化を進め、それぞれの地域の新人に移行したというものである。
「多地域起源説」を全面的に否定したのが分子人類学で、新しく「新人の単一起源説」「アフリカ起源説」が提唱された。曰く、現生人類はすべて二〇~一〇万年前にアフリカで生まれ、七~六万年前にアフリカを出て全世界に広まった。
現生人類の「アフリカ起源説」によれば、北京原人、ジャワ原人、ネアンデルタール人などの先行人類はすべて絶滅したらしい。このようなパラダイムシフトを起こしたのが、ミトコンドリアDNAの多様性に関する研究である。
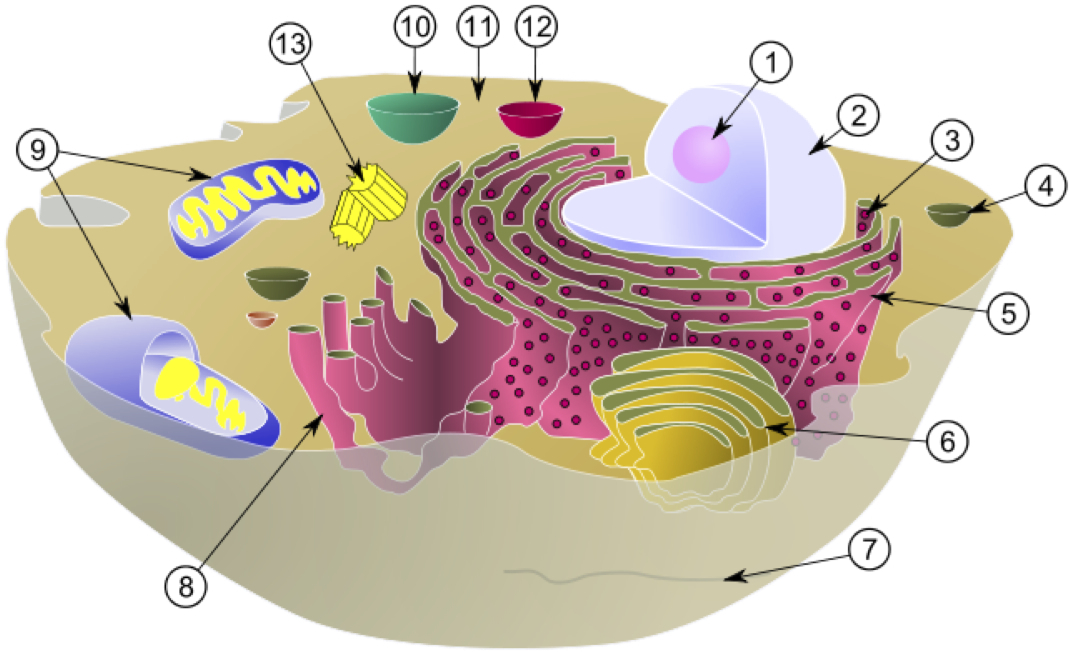
細胞の模式図
核 ② 以外にミトコンドリア ⑨ にもDNAが存在する
park6.wakwak.comより
DNAは核だけでなくミトコンドリアにも存在する。ミトコンドリアDNAはたいへん小さく、核のDNAの二〇万分の一の大きさしかない。約一六五〇〇〇個の塩基対が環状になったもので、核の染色体DNAに先行して全塩基配列が決定された。
ミトコンドリアDNAは母親から娘に受け継がれる。精子由来のミトコンドリアは受精時に徹底的に排除される仕組みがある。そもそも父親由来のミトコンドリアDNAは存在しない。
息子にもミトコンドリアDNAが受け継がれるが次の世代には行かない。娘経由のミトコンドリアDNAだけが娘へと代々受け継がれてゆく。逆向きに言うと、母親の母親のそのまた母親と、ずっと先祖を辿って行ったとしても、ミトコンドリアDNAは変わらないままである。
常染色体DNAは二本鎖で、代を重ねるごとに薄まってしまう。自分の常染色体DNAは片方が父親から、もう片方が母親から受け継がれる。父も母もそれぞれまたその両親から受け継いでいる。一世代前に比べると二分の一、二世代だと四分の一、三世代だと八分の一、n世代前からすると2のn乗分の1に薄まる計算になる。
しかしミトコンドリアDNAは違う。常染色体のようには代々薄まらない。長い時代にわたり、一定の配列を保つ。
だが、遺伝子にはランダムに点突然変異(mutation)が起こることがある。塩基がなくなったり(欠失=deletion)、塩基が入り込んだり(挿入=insertion)、他の塩基に変わったり(置換=substitution)する。数塩基にわたる欠損などの例もある。
母親に二人の娘がいると想定する。姉に受け継がれたミトコンドリアDNAのどこかに変異が起こり、妹には起きなかったとする。姉の子孫には代々その変異が受け継がれ、妹の子孫には母と全く同じタイプが代々受け継がれる。
代を重ねると他の部位でも変異が起こり、そのまた子孫に受け継がれる。こうして変異自体が蓄積されてゆく。上述の祖先の妹にも変異が起きたとしても、姉に起きた最初の変異と全く同じ部位で全く同じ塩基に置き換わったり、全く同じ部位で塩基が欠失したりする確率はほぼゼロと見なせる。
代々積み重なった変異のパターンができて来る。それを解析することにより、ミトコンドリアDNAはさまざまなグループに分けられる。そのパターンはハプログループと呼ばれる。ヒトのDNAはきわめて変異が起こりにくく、比較的容易に共通の祖先を捜すことができる。
二つのハプログループを取り出して繰り返して比較し、最も直近の共通祖先(MRCA=most recent common ancestor)を探す。得られた共通祖先どおしをつなげて遺伝系統樹を作成する。そうした解析によりアフリカの単一祖先にたどり着いた。
世界で最初に作られた系統樹が次の図である。
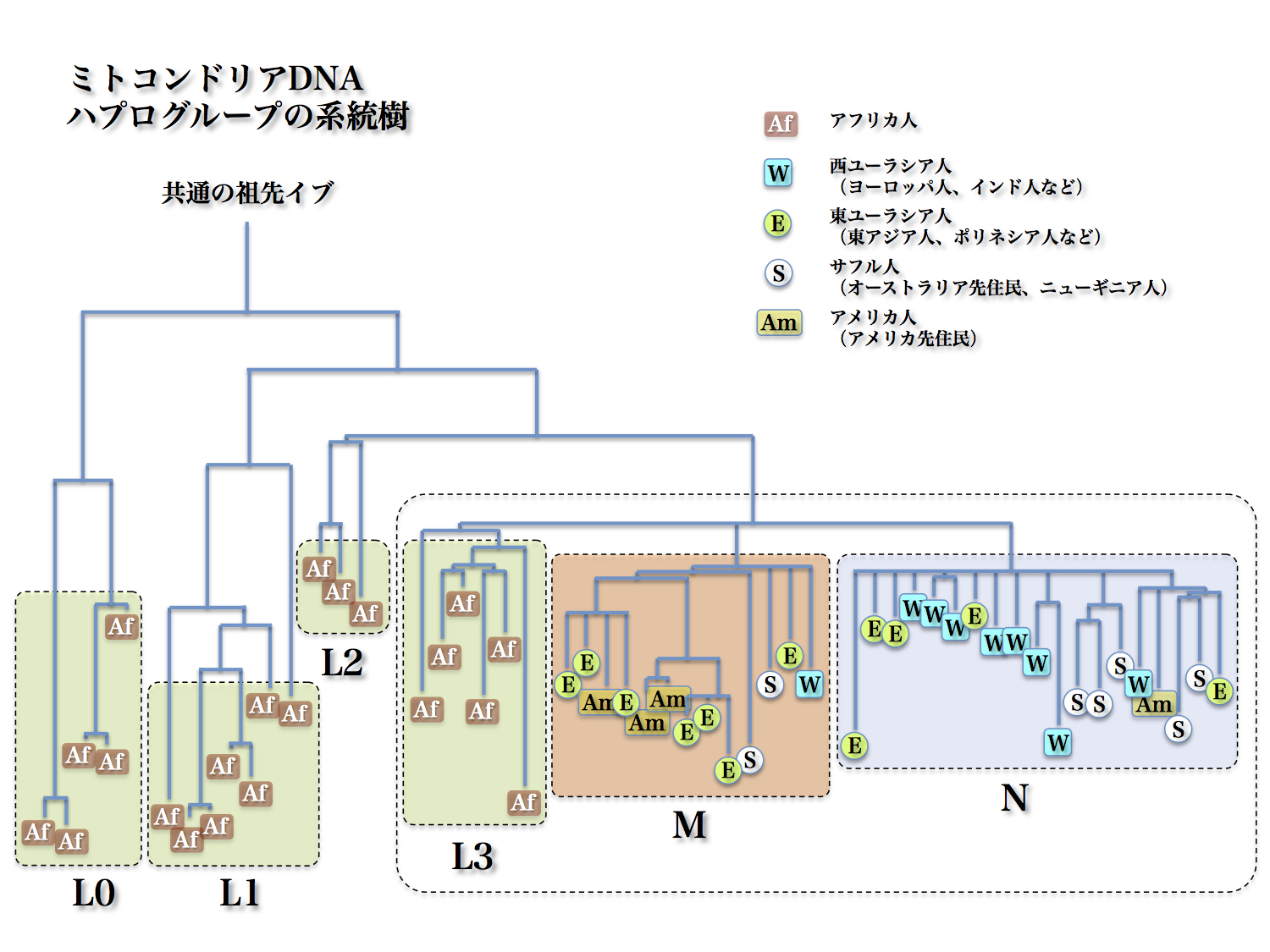
ミトコンドリアDNAハプログループの系統樹
篠田謙一著「日本人になった祖先たち」より改変
おおもとは Ingman et al. 2000 による。
図中の線の長さは遺伝的な距離を示している。
ミトコンドリアDNAハプログループを分類したところ、図のように大きく四つに分かれることが判明した。L0、L1、L2、そしてL3、M、Nを含む群(クラスター)である。アフリカにとどまったのがL0~L3クラスター、MとNは出アフリカを果たしたクラスターである。
世界に散らばった人々はMとNから出た。アフリカにいる人々は分子人類学的にはたいへん多様性に富んでいた。ヨーロッパ人とアジア人の違いよりも、アフリカ人どうしの隔たりの方が大きかった。
DNA解析という手法によって、ヒトは白人、黒人、黄色人種といった分け方とは全く違うグループに分けられることが判明したのだった。
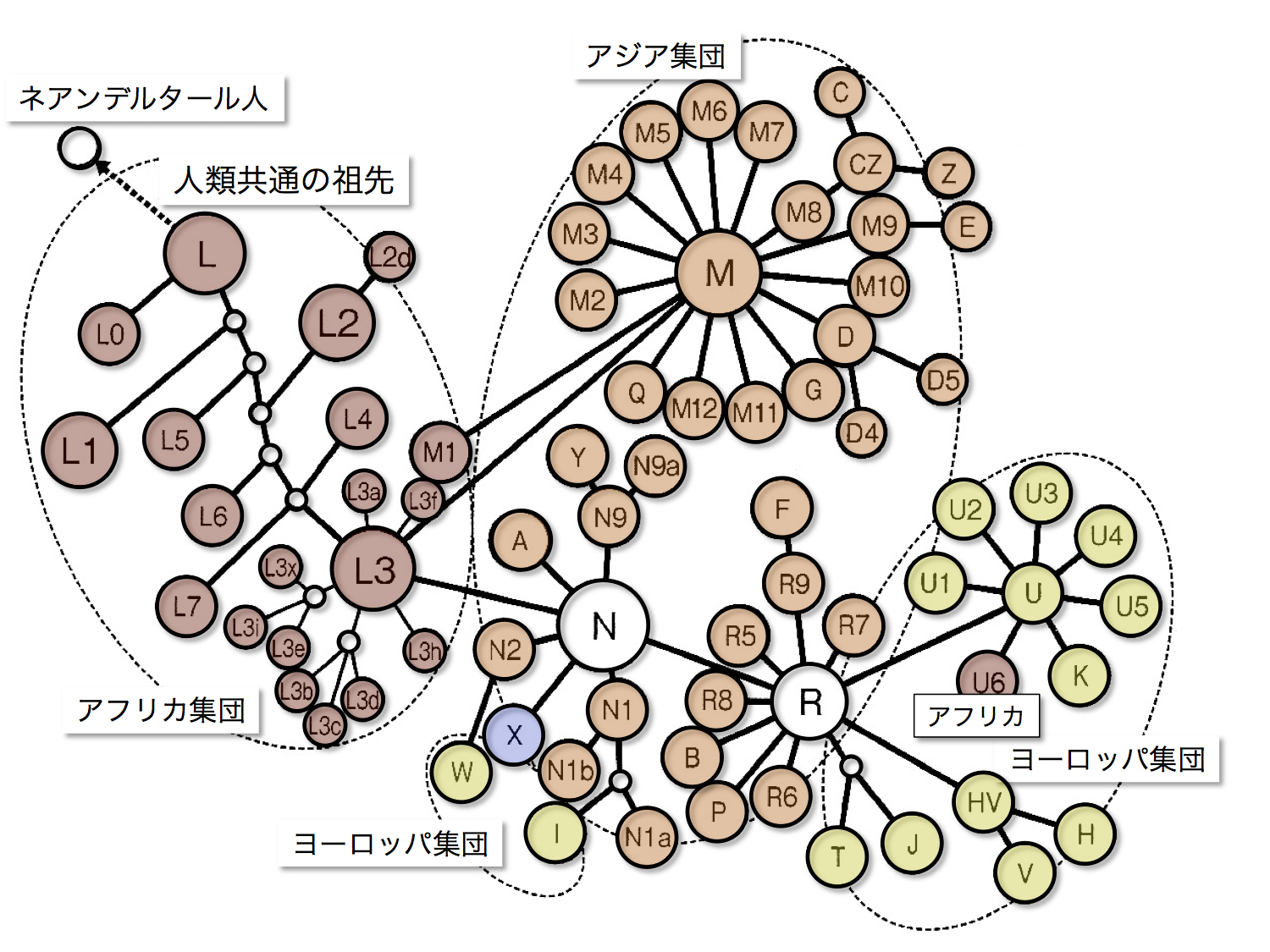
ミトコンドリアDNAハプログループ間の系統関係
篠田謙一著「日本人になった祖先たち」より
この図はミトコンドリアDNA各ハプログループの系統関係を示している。L(茶色)はアフリカで見出される集団、M、N、Rから派生するクラスターで構成されているアジア集団(オレンジ色)、主にRから派生したヨーロッパ集団(黄色)にまとめられている。
N由来のWと I はヨーロッパ集団に含まれ、同じくN由来のX(紫色)はアジアにもヨーロッパ集団にも存在する。Rから派生したUクラスターのうちU6だけはアフリカに出戻った。MクラスターのM1も同様にアフリカに存在する。
アフリカのL0~L3は混み入っていて星型をほとんど示さない。対してMはたいへんきれいな星型を示し、Nやそれから派生するR、そこからまた派生するUのクラスターも星状である。
一五世紀終わりから一六世紀に始まる植民地時代、ヨーロッパから新大陸へ、アフリカから新大陸へ、人類に大規模な移動が起こった。だが分子人類学ではそれより以前、つまり先史時代の人類の移動、拡散と分布を主な研究対象にしている。
現在、さらに多くのサブハプログループが報告されている。2013年度の最新系統樹詳細はここを参照されたい。
Ⅱ)世界各地域におけるミトコンドリアDNAハプログループ構成頻度
まずは全世界におけるミトコンドリアDNAハプログループの分布頻度を概観する作業から始める。
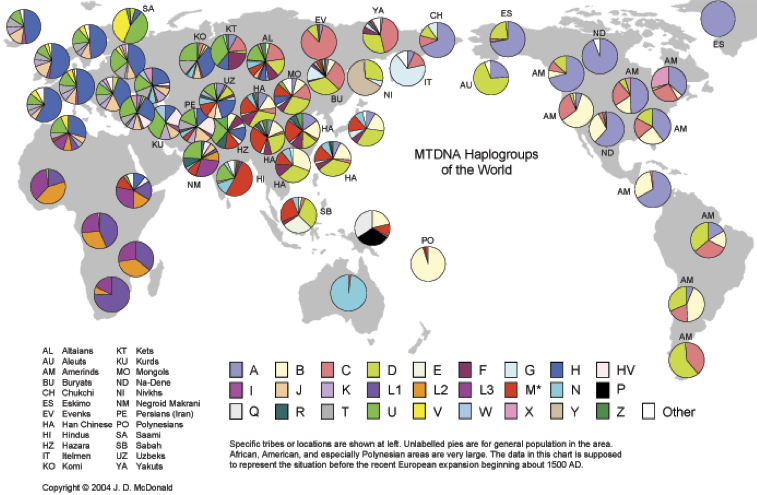
ミトコンドリアDNAハプログループの世界分布
http://www.scs.illinois.edu/~mcdonald/WorldHaplogroupsMaps.pdf より
さらに大きな図はこちら
ⅰ)アフリカ大陸
サハラ以南ではL1、L2、L3がほとんどすべてを占める。エチオピアではH、M、HV、Uなどが加わり多彩な構成になっている。サハラ以北ではH、U二つが半数弱、L1、L2、L3の三つで25%と、ヨーロッパの構成にサハラ以南の構成が混じる形である。
ⅱ)ヨーロッパ
半数前後をHが占め、J、K、T、U、Vなどが混じる。極北のサーミ族(Saami)ではU、Vで八割と特徴的である。
ⅲ)西アジア
トルコ、コーカサスではH、Uがそれぞれ1/5から1/4で、ほかT、J、Xなどが続く。クルド人はUが三割以上、HVがそれに次ぎ、H、J、K、Wが均等に混じる。ペルシャ人はもっと多彩で、HV、J、U、Hで七割を占め、T、K、そしてM、Nなどが続く。
ⅳ)南アジア
インドではMが半数を占める。次いでU、Nの順。それらで約八割となる。
ⅴ)東南アジア、華南、華北
ほとんどはD、B、M、F、N、Aである。それぞれの分布傾向の違いは後述する。
ⅵ)日本
別途詳述するが、基本的に東アジアでよく似た構成となっている。
ⅶ)モンゴル
筆頭はD、次いでC、M、H、B、F、Aの順に多く、以上で約八割に達する。
ⅷ)中央アジア
ウズベキスタン人はH、J、Uなど欧州系のハプログループとM、A、Dなど東アジア系が混じっている。アフガニスタンにいるイラン系のハザール人のデータも載っている。U、Z、H、M、F、B、Rの順の頻度で、R、Zの存在が特徴的だろう。
ⅸ)北東アジア
エヴェンキ(EV)はC、ブリヤート(BU)とヤクート(YA)はCとD、ニヴフ(NI、Nivkh)はYに次いでD、イテリメン(IT)はG、チュクチ(CH)はAが特徴的。
ⅹ)アメリカ大陸
アリュート(AU)はD、エスキモー(ES)と極北に住むナ・デネ語族(ND)がA、北米西部はBと次のC、北米中西部はAを筆頭にCとB、D、現在のカナダ東部先住民はA、C、X、メキシコ国境の米先住民と中米先住民はA、Bが特徴的である。南米ではB、C、D、Aの混じりで、南下するとCとDでほとんどすべてとなる。
Ydnaの場合がほとんど単一のハプログループであるのに対し(後述)、ミトコンドリアDNAのハプログループは多様性が保たれている傾向がある。
ⅺ)オセアニア
ニューギニアがP、Q、B、Mと多彩であるのに対し、オーストラリアではNがほとんどすべてで、ポリネシアではBが大部分となっている。
Ⅲ)ミトコンドリアDNAハプログループからみた人類の推定移動拡散経路
続いて、ミトコンドリアDNAハプログループがどのような経路で、世界のどの地域に移動拡散していったのであろうか。提唱されている推定経路を紹介する。
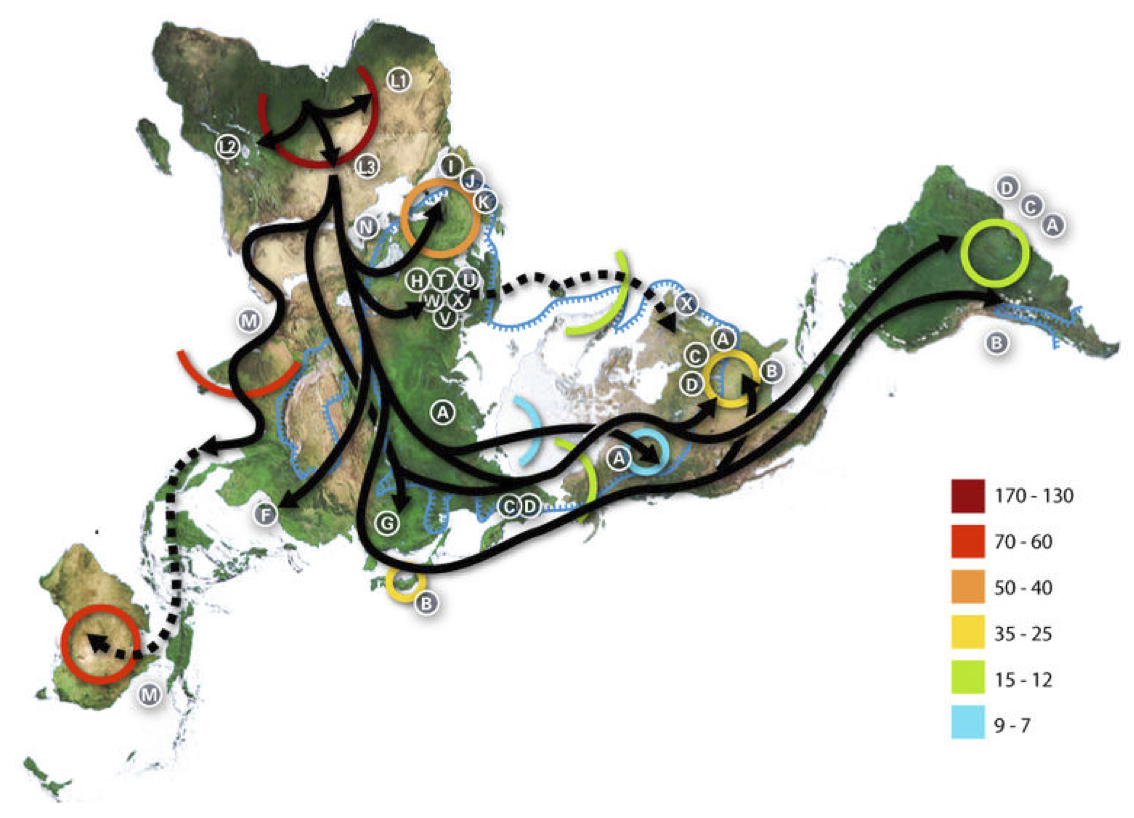
イブの子孫の出アフリカと世界各地への移住
Wikipediaによる(CC BY-SA 3.0)
より大きな図はこちら
ダイマクション地図のアニメーションと展開図はこちらを参照されたい。
この図はイブの子孫がアフリカを出たあと、どのような経路をたどってどの地域に移動したか、主なハプログループの動きを示している。
図に描かれていることは、主に次の通りである。
1)ハプログループMがアラビア半島、インド大陸、東南アジアを経て
オーストラリアまで行った。
2)M由来のGが東アジアに、CとDが東アジアやアメリカ大陸に移動し
た。
3)NからRを経てF、Bが分かれ、日本を含む東アジアに移り住んだ。
4)Bはさらに旅を続け、N由来のAもアメリカ大陸にまで移った。
5)N由来の I 、X、W、ならびにRから派生したJ、K、U、H、V、Tが
ヨーロッパ人の母となった。
色付きの円、弧は、何千年前という年代を表している。たとえばL1~3クラスターがアフリカで拡がったのが一七~一三万年前、Mがインドに定住したのが七~六万年前、Bが日本列島に行き着いたのが三万五千年前から二万五千年前頃など。
北米大陸にA、C、B、Dが住み着いたのも三万五千年前から二万五千年前、A、C、Dが南米大陸に行き着いたのが一万五千年前から一万二千年前、I 、J、Kがヨーロッパに展開したのが五~四万年前などと大まかなことが描かれている。
残念ながらアジアへ移り住んだ集団については、日本列島への到達に関する詳細も含めて本図には描かれていない。
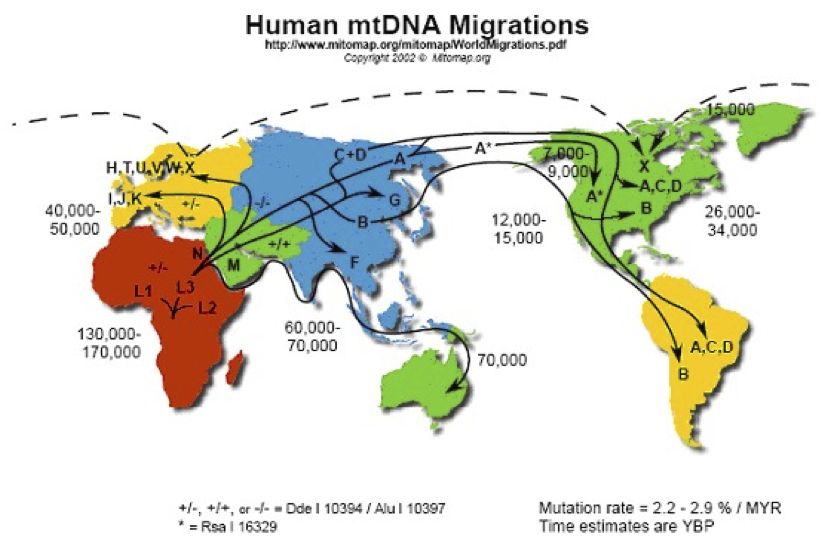
ミトコンドリアDNAの世界各地への旅
http://www.mitomap.org/mitomap/WorldMigrations.pdfより
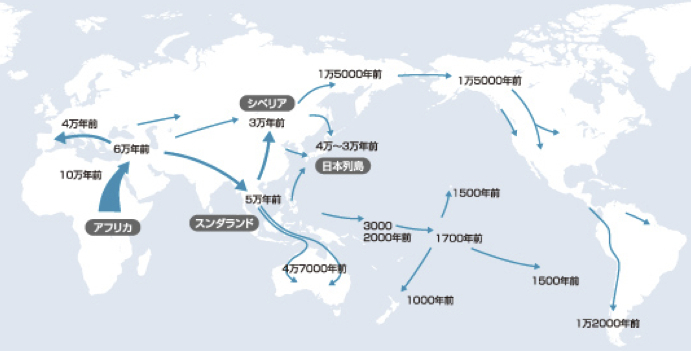
ミトコンドリアDNAハプログループから推定される人類の世界拡散経路とその時期
篠田謙一著「日本人になった祖先たち」より改変
これら二つの図は、見慣れた形の世界地図に落とし込まれた、イブの子孫たちの旅路である。年代や経路については多少の相違があるものの、世界各地への人類拡散の推定経路とその年代が書き込まれている。
日本列島には北から、朝鮮半島経由でシナ大陸から、南から人々がたどり着いたというイメージで描かれている。四~三万年前のことだという。太平洋沿岸の島嶼(とうしょ)に展開したのが三万~一万五千年前、イースター島やニュージーランドへの移住が一五〇〇~一〇〇〇年前と推定されている。
ただこうした移動経路は、現在人のミトコンドリアDNAハプログループの解析から推定したものである。考古学的な資料を使って導き出されたものではない。
サイズの小さなミトコンドリアDNAは現代人の生きたサンプルばかりでなく、条件が良ければ考古学的な試料からも解析可能である。今後はそうしたサンプルからの解析が積み重ねられ、研究が進展してゆくことが望まれる。
Ⅳ)日本人のミトコンドリアDNAハプログループ
ⅰ)日本人のミトコンドリアDNAハプログループ構成頻度
次に、日本人のミトコンドリアDNAハプログループについて紹介する。
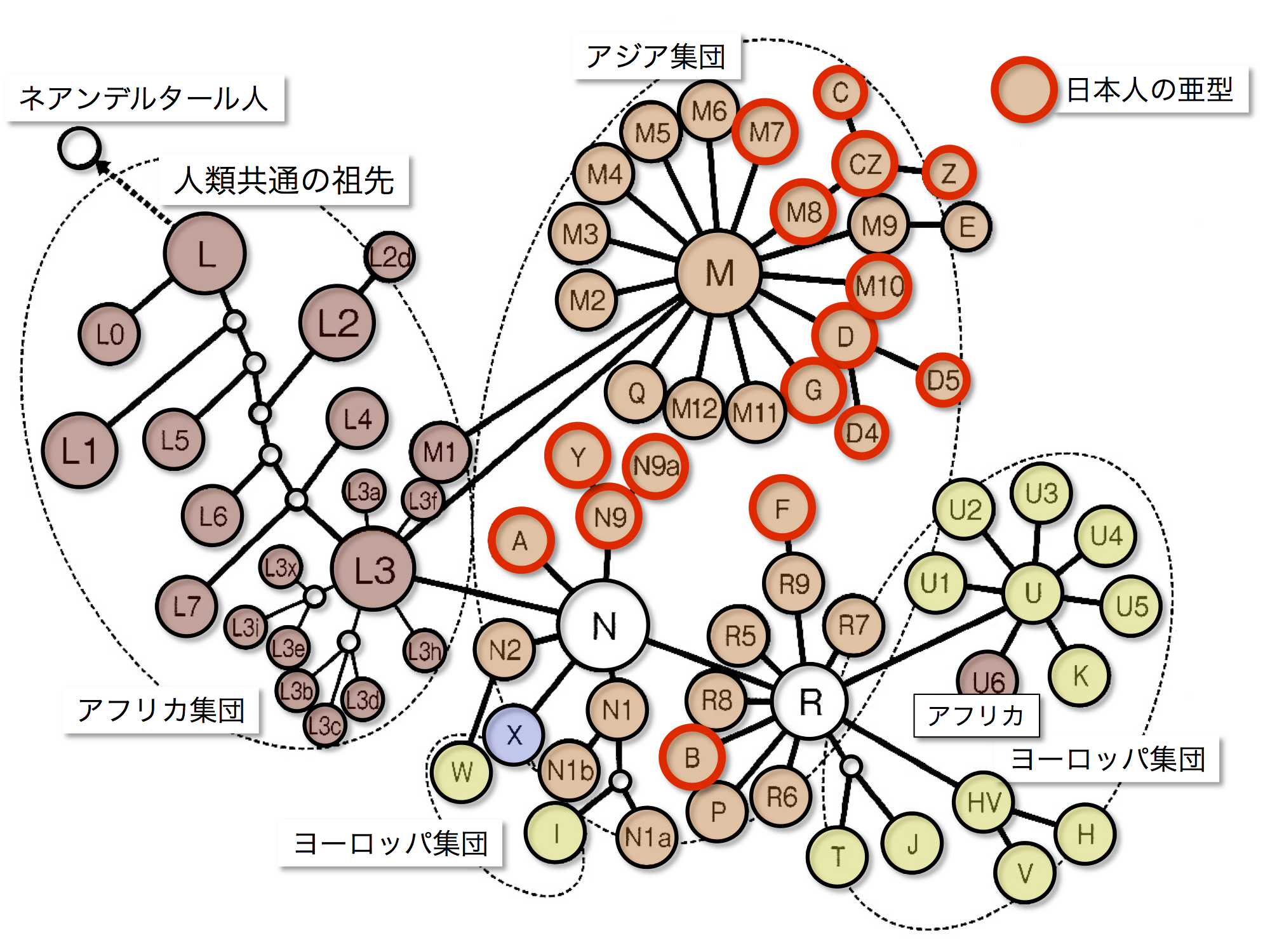
日本人に存在するミトコンドリアDNAハプログループ
篠田謙一著「日本人になった祖先たち」より改変
ミトコンドリアDNA各ハプログループの系統関係の図に、日本人に認められるハプログループを赤丸で示した。日本人の主なハプログループはM、N、Rいずれかのクラスターに属している。
Mクラスター:M7、M8、M10、M8に由来するCとZ、D4、D5、G。
Nクラスター:A、N9由来のY、N9a。
Rクラスター:B、R9由来のF。
図には示されていないが、次のサブハプログループも日本列島に一定の割合で見出されている。N9由来のN9b、B由来のB4、B5、M7に由来するM7a、M7b、M7c、M8由来のM8aである。
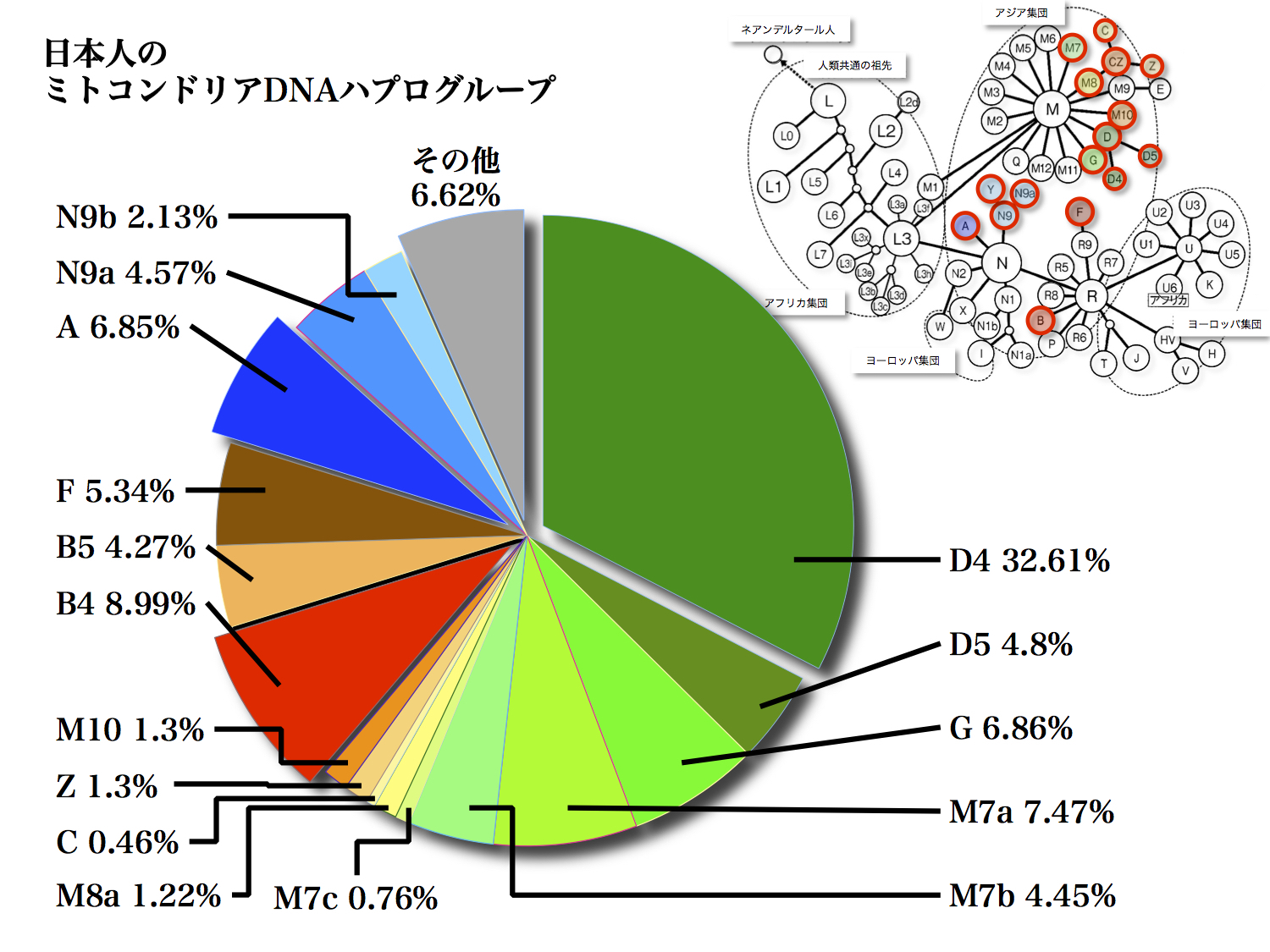
日本人のミトコンドリアDNAハプログループの頻度
篠田謙一著「日本人になった祖先たち」より改変
この図は日本人のミトコンドリアDNAハプログループの頻度を円グラフで表している。緑色系統から黄色を経てオレンジ色に至るのがMクラスター、赤から茶色に至るのがRクラスター、青系統がNクラスターにそれぞれ属するハプログループ、サブハプログループである。
頻度2%以上のハプログループは次の11種類である。
D4、D5、G、M7a、M7b、B4、B5、F、A、N9a、N9b。
これらハプログループの発端になった女性たちこそ、代表的日本人の母11人と呼ぶこともできるだろう。篠田氏は、2%未満のM7c、M8a、C、Z、M10も加えて16のルーツとしている。次からそれぞれ概説を紹介しよう。
ⅱ)ミトコンドリアDNA各ハプログループの概説
<Mクラスター>
<D>
日本人第一のグループで37.4%を占める。起源は35000年前に遡る。南回りで東アジアに入ったハプログループMの中から、最終氷河期の最寒気少し前に誕生した。
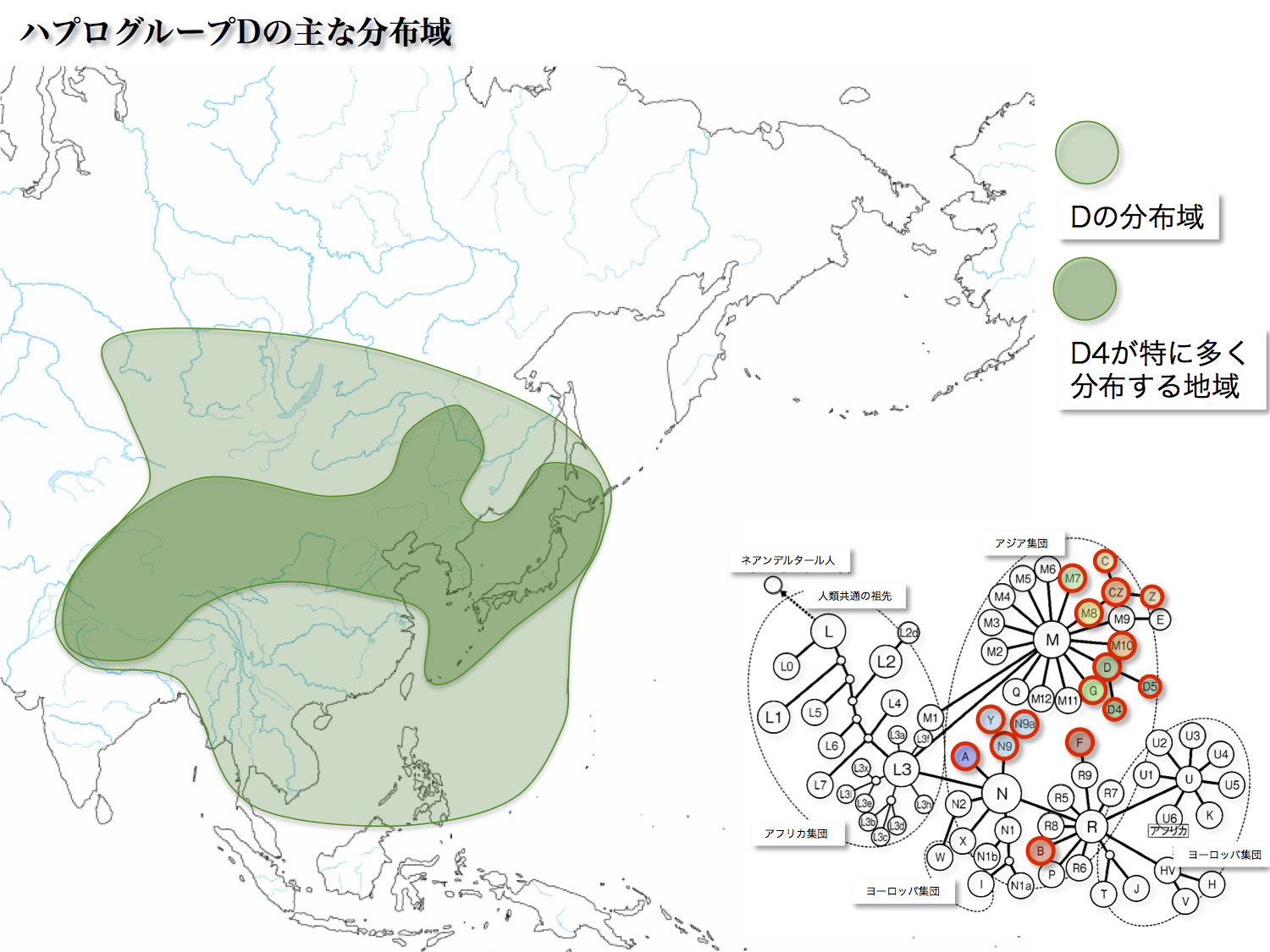
ハプログループDにはD1~D6のサブハプログループが存在する。D1とD2はアメリカ先住民に見出される。D4とD5が中央アジアから東アジアにかけて最も優勢に分布しており、特に日本列島、朝鮮半島、中国東北部で三割から四割に達する。
サブハプログループD4はさらにD4a~D4nに分かれ、日本を含む東アジア東北部を中心に分布している。他方D5はD5a~D5bに枝分かれし、華南を中心とした分布となっている。
対照的にD4は日本で32.6%、華南で9.6%、D5は日本で4.8%、華南で4.8%という比率である。D4がいかに日本列島で多いかがわかる。
<G>
日本人の6.86%にあたる。最終氷河期最寒期以降。カムチャッカ半島や北シベリア先住民族に3~7割という高頻度で認められる。なぜそれほどまでに高頻度なのかという説明として、ボトルネック効果と言って、もともとは存在していた他のハプログループが消失したため、偏った亜型頻度を持つようになったとされている。
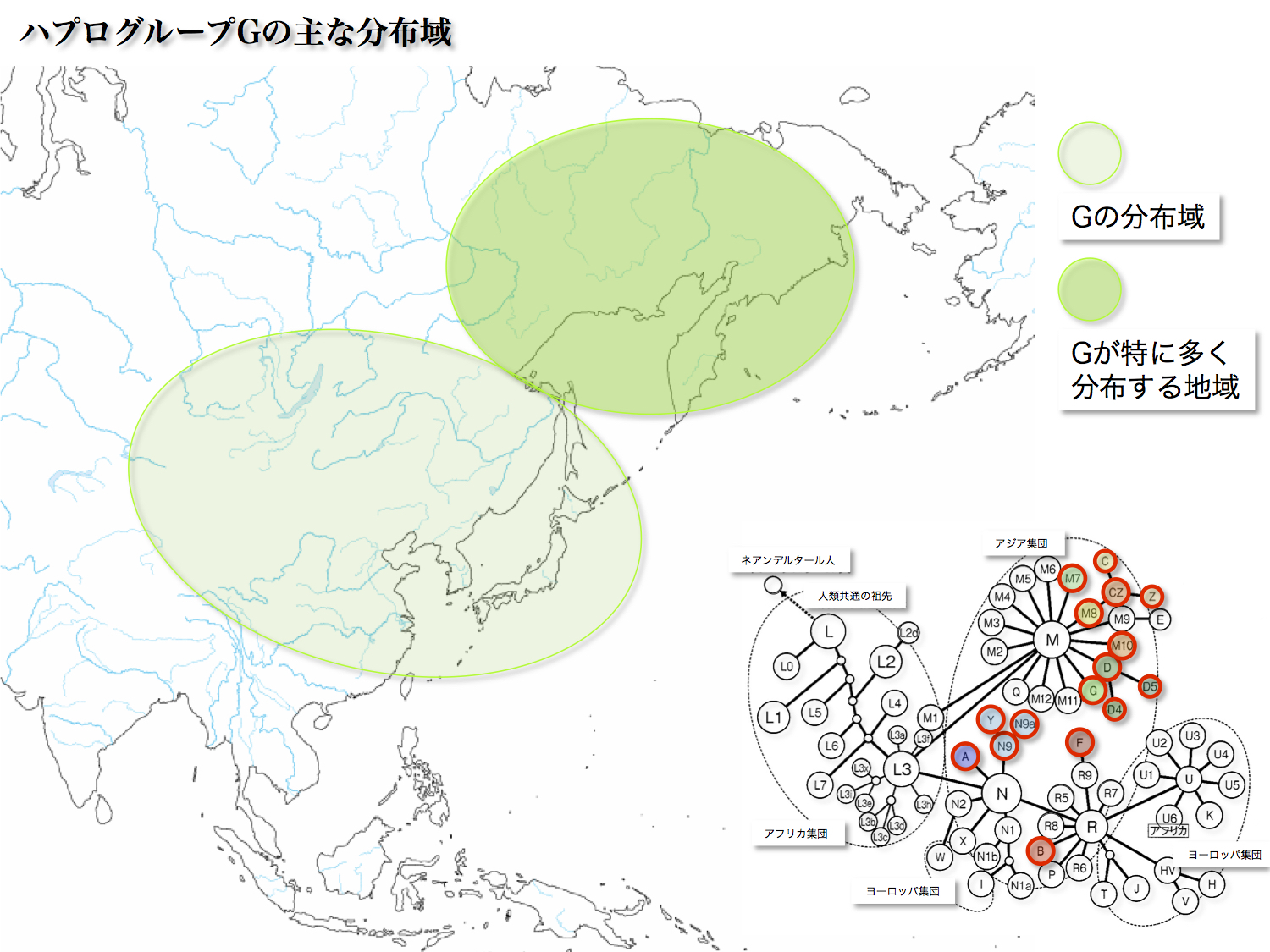
ハプログループGはG1~G4のサブハプログループに分類される。G1は日本本土、アイヌに分布し、朝鮮半島にも少数見られる。G2は中央アジアに存在し、東南アジア、華南にはほとんど分布していない。あまりハッキリとした分布境界を持たないものの、G3は中央アジア、モンゴルで見られる。
<M7>
ハプログループMはM7a、M7b、M7cのサブハプログループからなる。起源は40000年前、各サブハプログループは25000年前と推定される。海面が低下していた氷河時代に陸地だった部分が東南アジアにはあった。そこに住み着いた人々から分かれた集団が、それぞれの場所に定住したのではないかと考えられている。
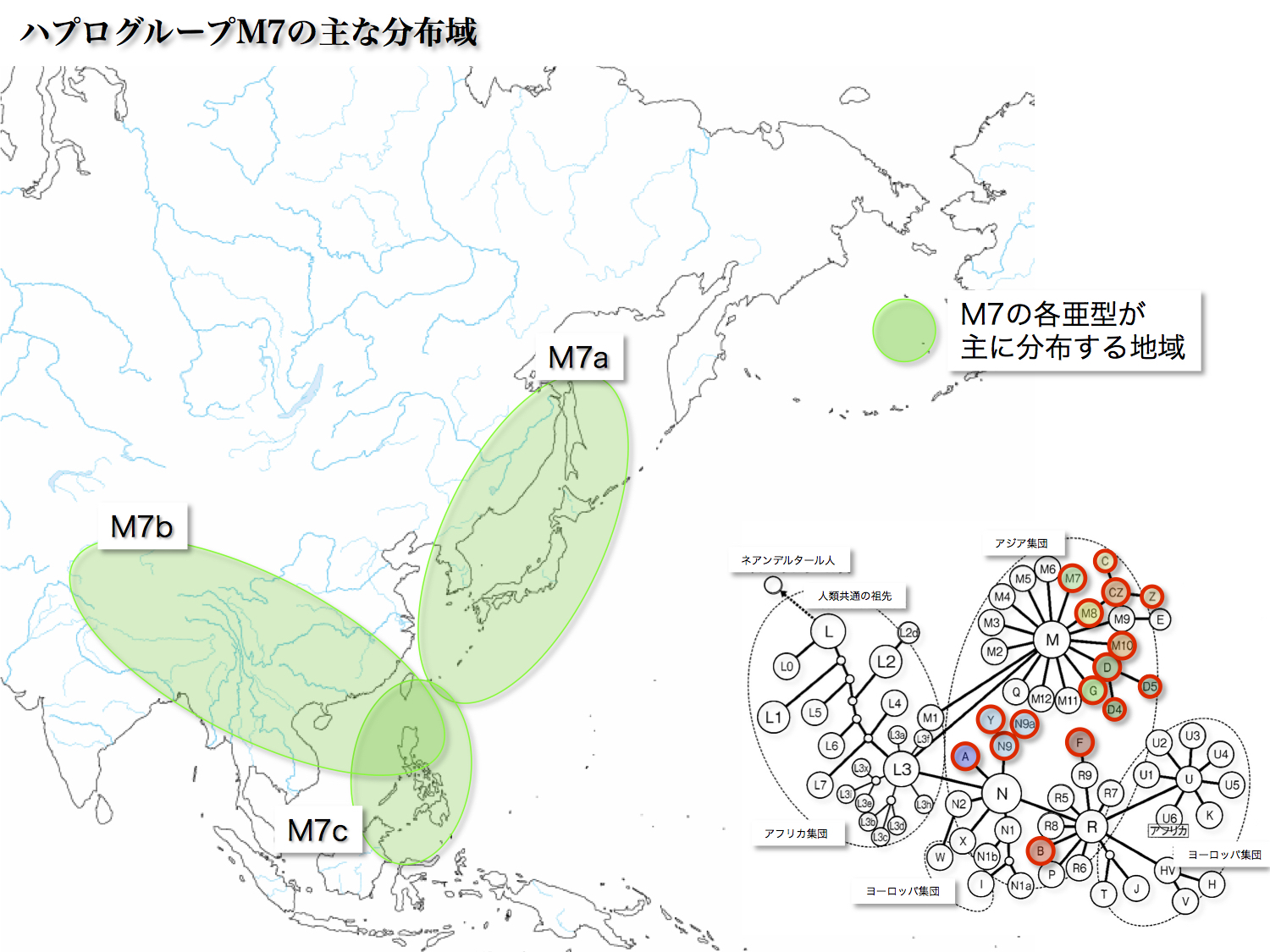
M7aは主に日本列島に存在し(7.47%)、特に沖縄に多く分布する(24.2%)。
M7bはシナ大陸沿岸から華南にかけて多い。日本列島だと4.45%の頻度である。
M7cは東南アジア島嶼部に多く認められ(最大18.8%)、日本列島だと0.76%と少数である。
いずれも中央アジアや北東アジアにはほとんど分布していない。
<M8>
ハプログループMはM8a、C、Zのサブハプログループからなる。
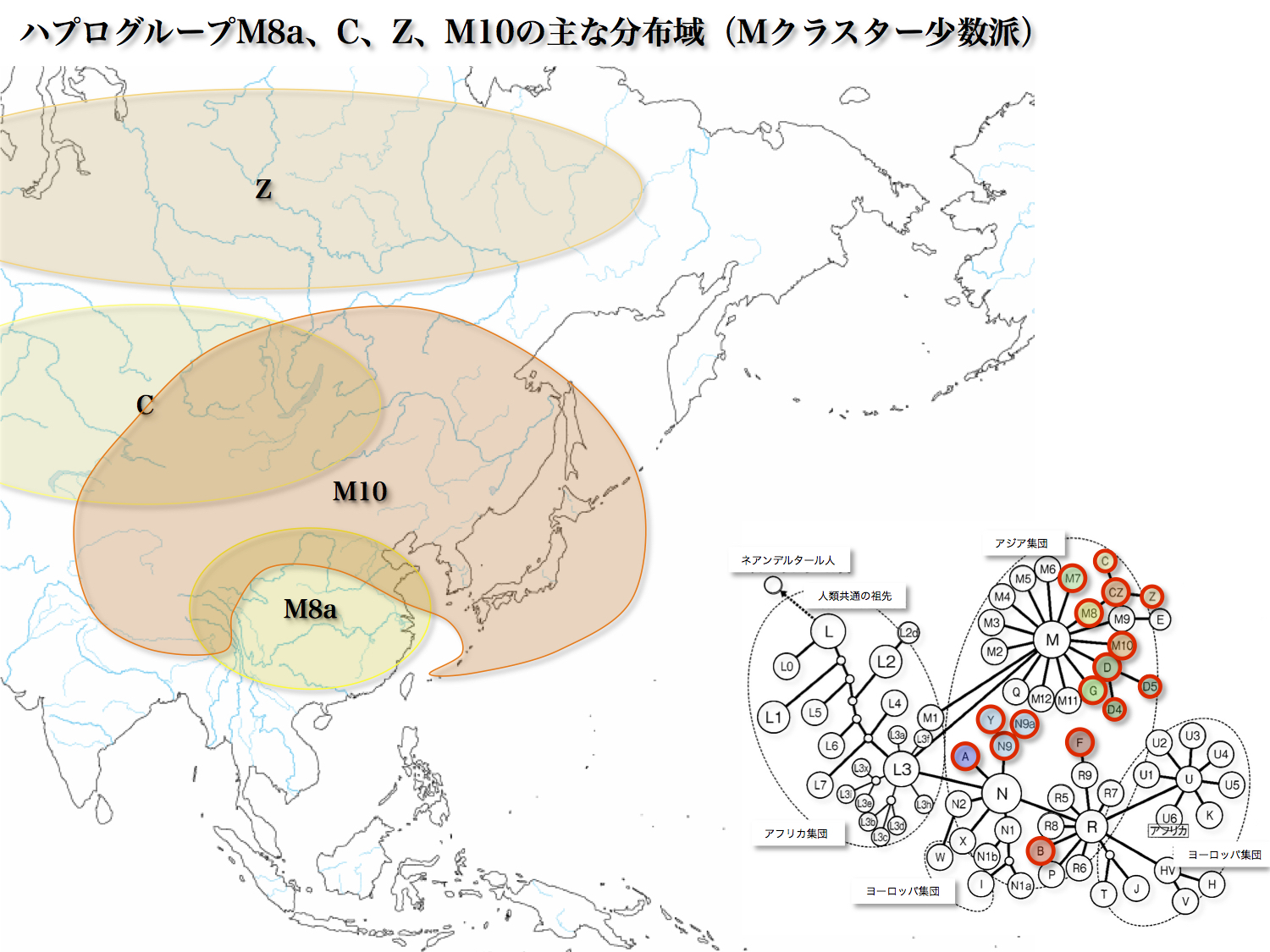
サブハプログループM8aは日本人の1.2%に相当する。華南と華北にも一定割合で出現し、周辺部には少ない傾向がある。
ハプログループCは日本人の0.5%に認められ、中心アジアから新大陸にかけて分布する。起源は30000年前とされる。朝鮮半島、華北、中心アジアの集団に大きなグループ内変異がある。中心アジアの地域集団が、遊牧民として勢力を拡げた時にそのテリトリーを拡げただろう。例はモンゴル民族である。
ハプログループZは日本人の1.3%にあり、他にカムチャッカ半島、フィンランド、朝鮮半島などに分布している。ヨーロッパと極東アジアにまたがる分布域を持っている特別なサブハプログループと言える。
<M10>
日本人の1.3%がM10であり、チベットでは8%を占め、ほか華北、ブリヤート、モンゴル、中央アジア、朝鮮半島に存在する。北の回廊を形成している亜型ではないかとも言われている。
<Rクラスター>
<B>
日本人第二のグループで13.26%を占める9塩基対欠損変異である。起源は40000年前で、インドから東南アジアに拡散したR集団の一つから誕生した。華南、東南アジア、南米山岳地域に分布している。
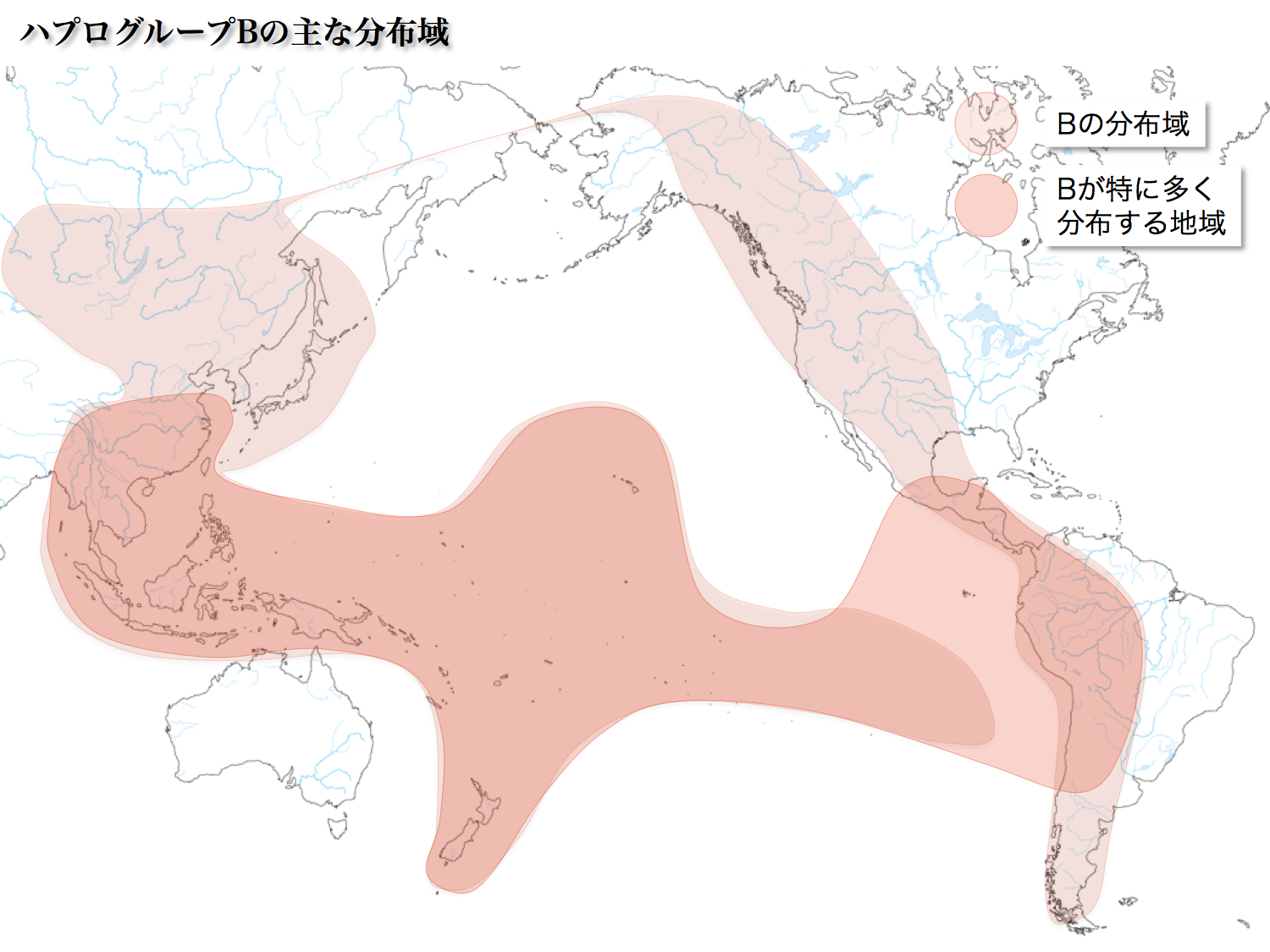
北米、南米には15000~12000年前に、南太平洋には6000年から1000年前頃に展開していった。B4、B5、B7(B6)がサブハプログループとして存在している。
太平洋島嶼に漕ぎ出したBの集団が、東進し南米にたどり着いたという仮説が出されている。中南米先住民にBの比率が著しく高い(24~63%)のが理由だ。北米では比率が8~9%と低い。
ベーリング海峡を経て北回りで到達した北米先住民のB集団は、その後中南米にまで足を伸ばし定住していた。ずっと時代が下って、太平洋からやってきた同じB集団の人々と遭遇したというのである。ロマンを誘う。ただ男性には厳しい現実が待っていたかもしれない。後述する。
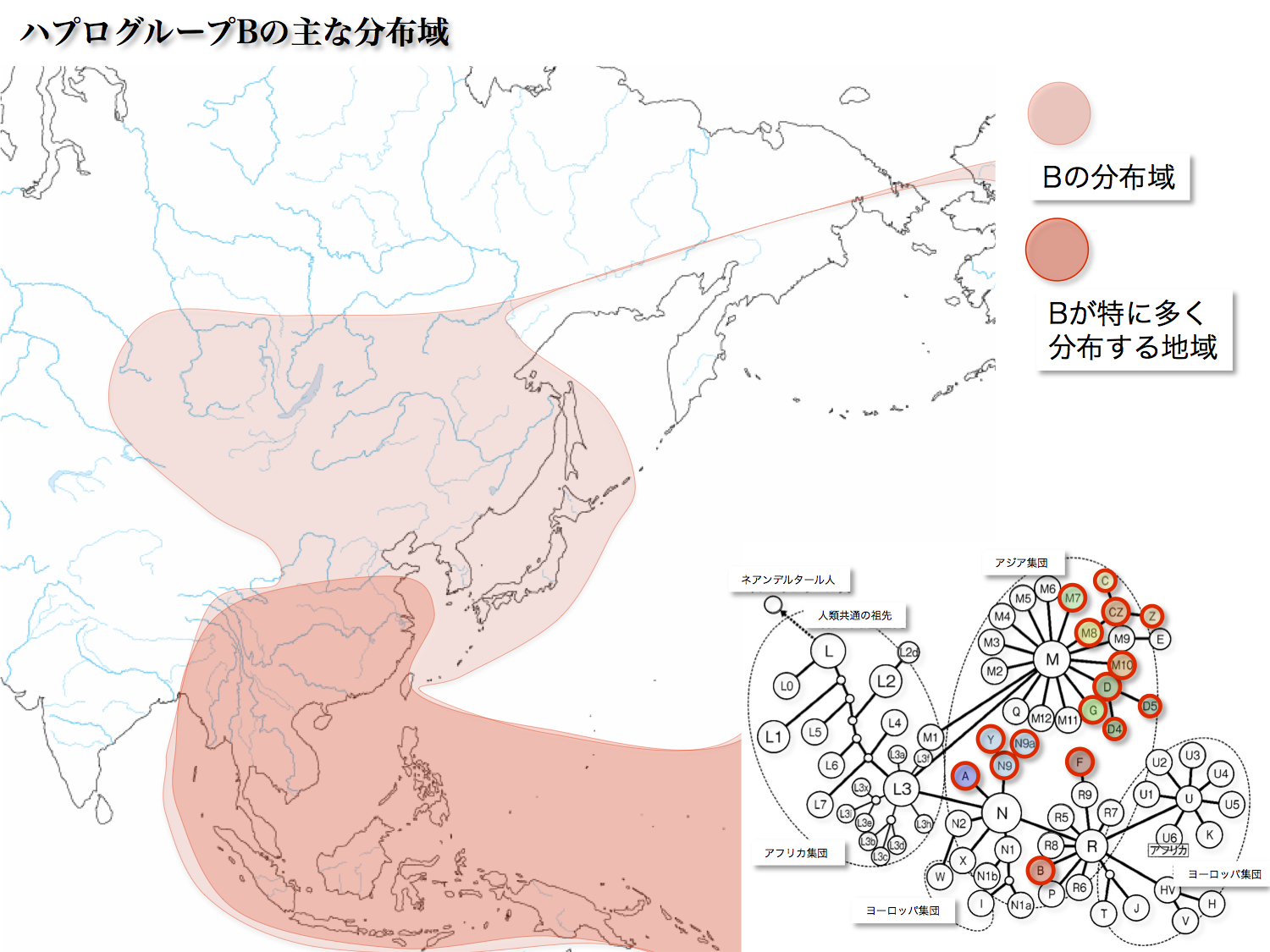
サブハプログループB4、B5については、次のように推定されている。東南アジアに来たR集団の一つから発生したBが、華南に進出したり、日本列島を経由して北米、南米に移動したり、大きく移動拡散していった。その経由地の日本にとどまり日本人の祖先となった女性たちがいたというのである。
<F>
日本人の5.34%を占める。起源は40000年前で、東南アジアに分布の中心(頻度として約12~37%)がある。
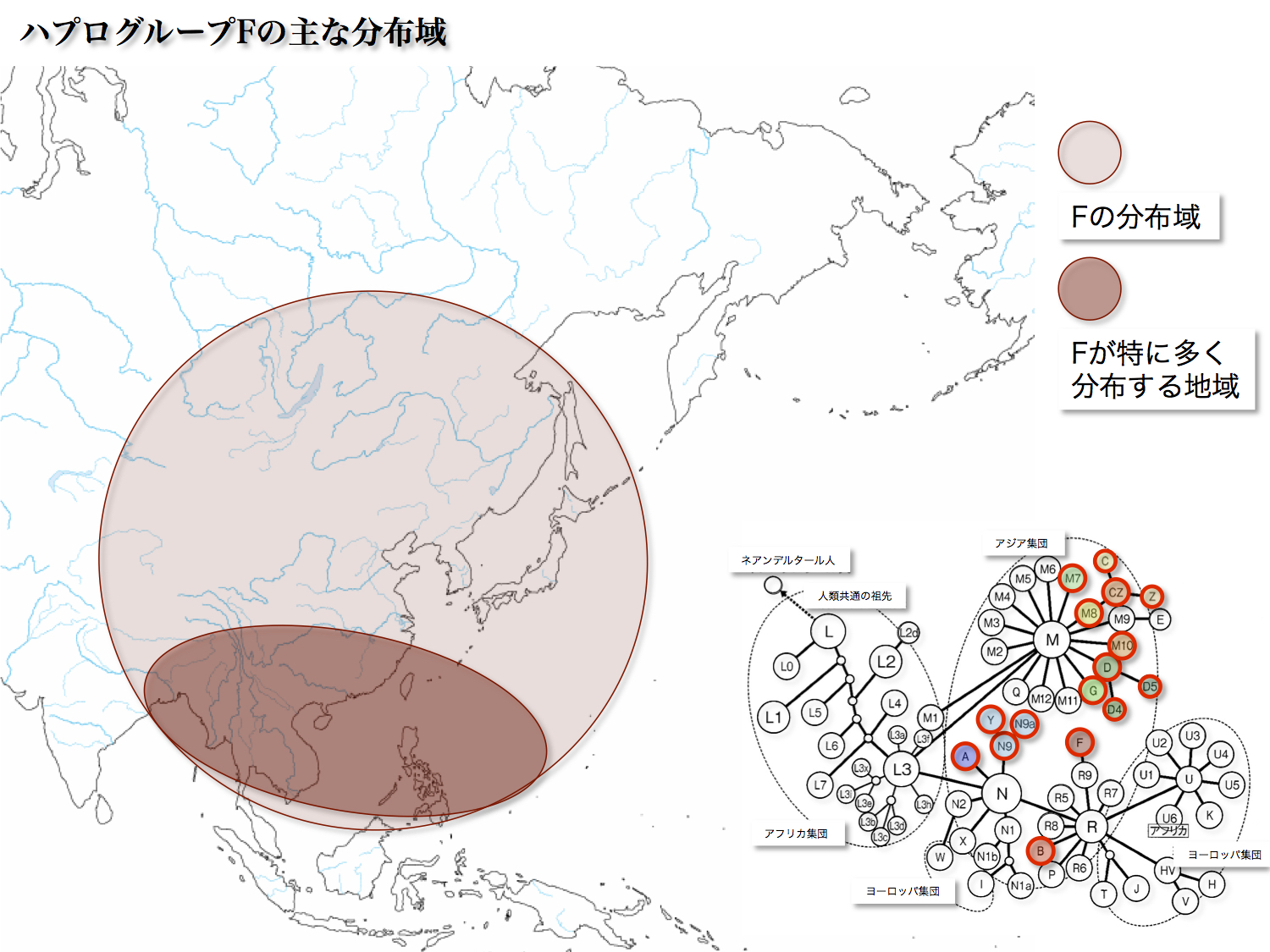
ハプログループRから分離しているところはハプログループBに似ている。しかしBとはちがって、アメリカ大陸にも南太平洋の島嶼への展開しなかった。FのサブハプログループにはF1、F2、F3、F4の四つがある。
<Nクラスター>
<A>
日本人の6.85%を占める。起源は30000年前、バイカル湖周辺とされる。マンモスハンターと呼ばれる集団の多数を占めていたと推定されている。
A3、A4、A5、A7、A8、A9のサブハプログループが報告されている。以前のA2は現在のA4に含まれる。
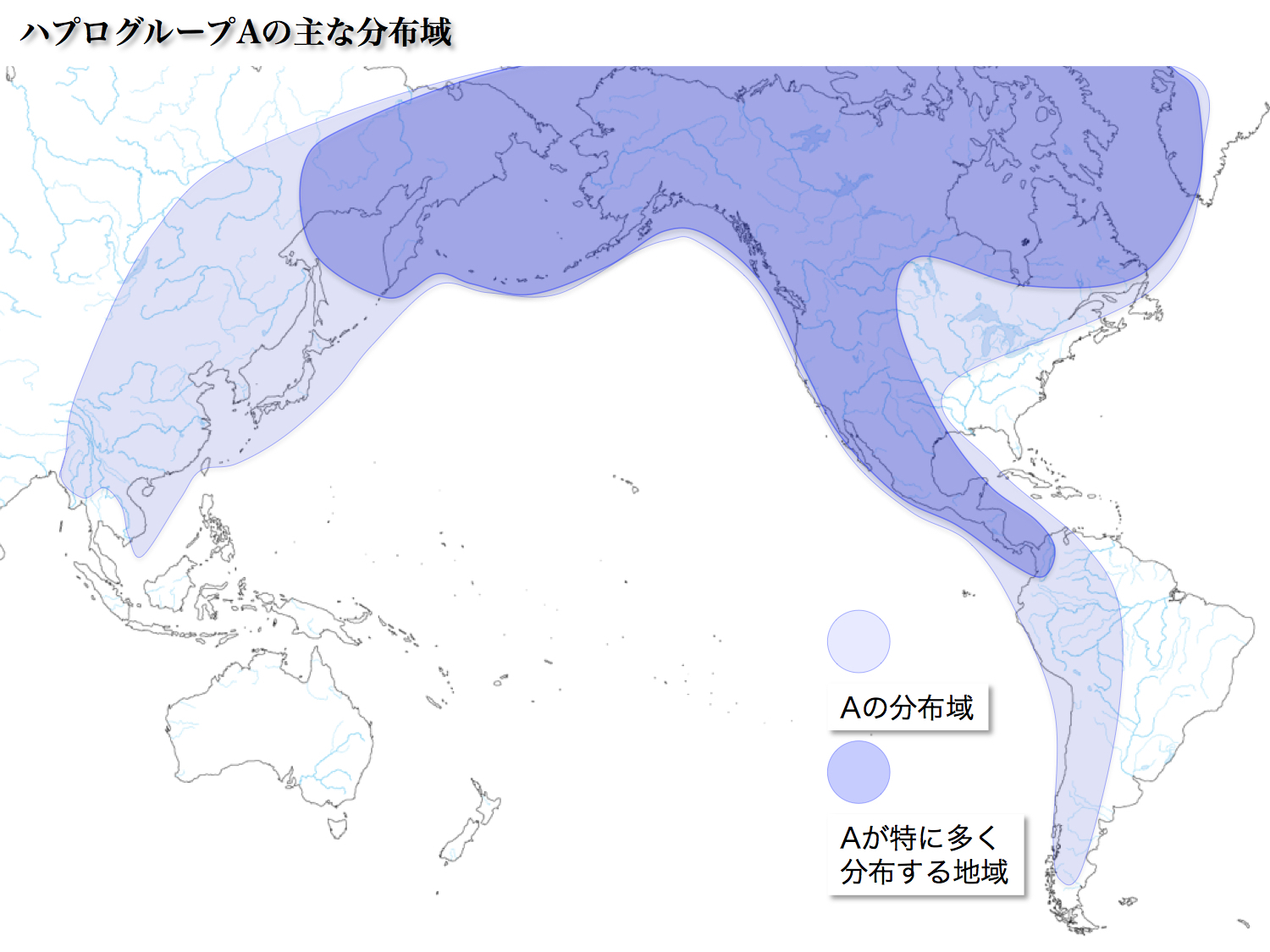
以前のA2がアメリカ大陸先住民に見出された。A4は東アジア全域に見られるが、A5は朝鮮半島、日本列島に分布が限られる珍しいタイプである。A5は7000年プラスマイナス2800年前に分岐したとされ、比較的新しいサブハプログループである。
<N9>
ハプログループN9は、N9a、N9b、Yというサブハプログループに分かれる。
ハプログループN9は、Rを経由せずにヨーロッパに入ったW、 I 、アジアとヨーロッパに広がったXとともに中東から北方に進み、そこで別れてヒマラヤの北を通って東アジアへ拡散したと推定されていた。
現在ではAとともにインドを経由して東南アジアに入った集団Nの中にN9がいて、東南アジアから東アジアに展開したのではないか言われている。本稿最終章を参照されたい。
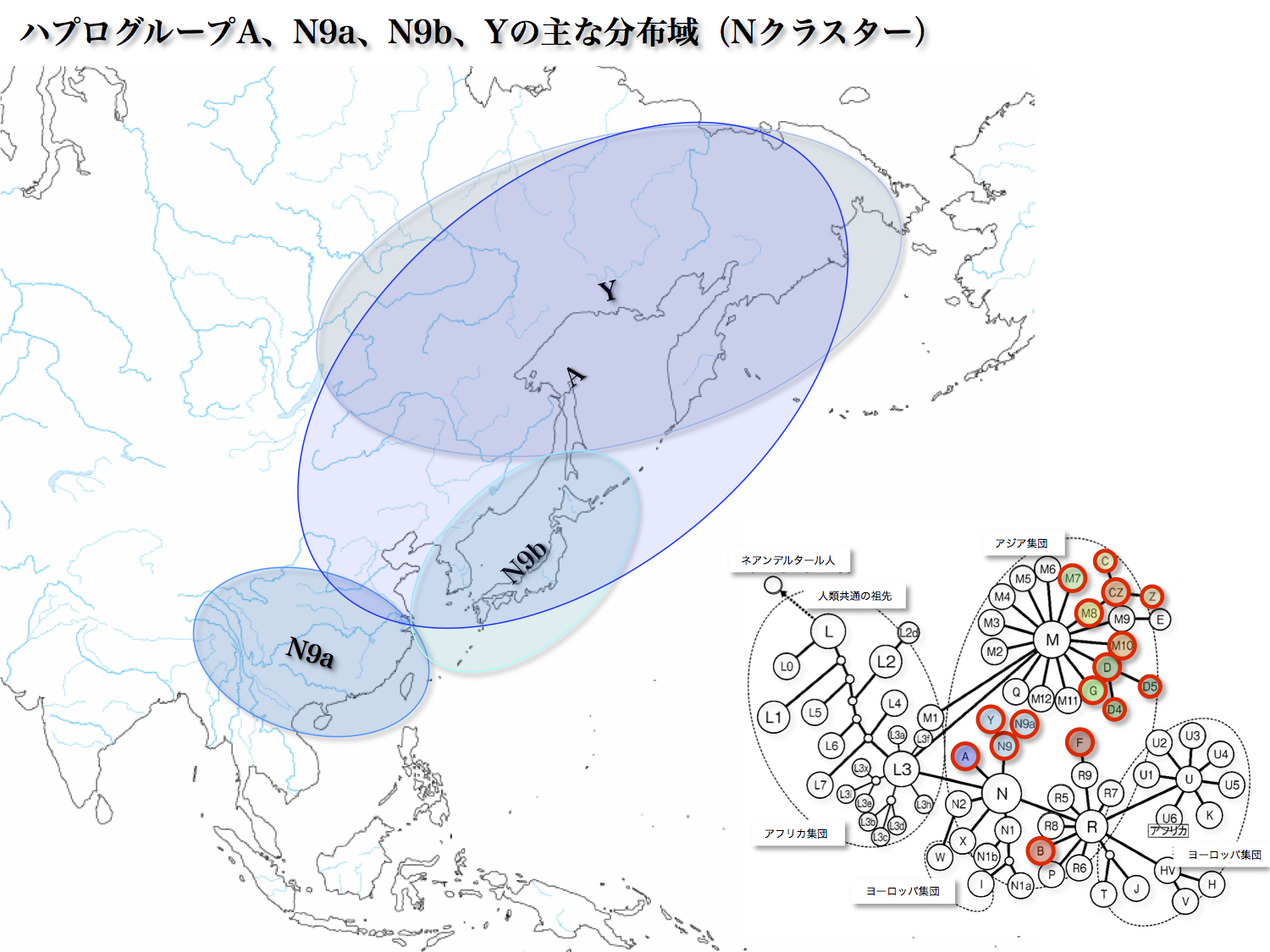
サブハプログループN9aは日本で4.57%存在する。東アジア~東南アジアの広くに分布し、華南や台湾先住民に比較的多いという。
サブハプログループN9bはほぼ日本列島にのみ(2.13%)存在し、朝鮮半島や沿海州にわずか分布する。北方ルートの代表ではないかと言われている。
サブハプログループYは北東シベリアにだけ多く、アイヌにも存在する。日本ではわずか0.4%だけである。
ⅲ)日本人に特徴的なM7aとN9bについて
日本人に特徴的なハプログループとしてM7aとN9bがあげられる。ほぼ日本列島にのみ分布する。
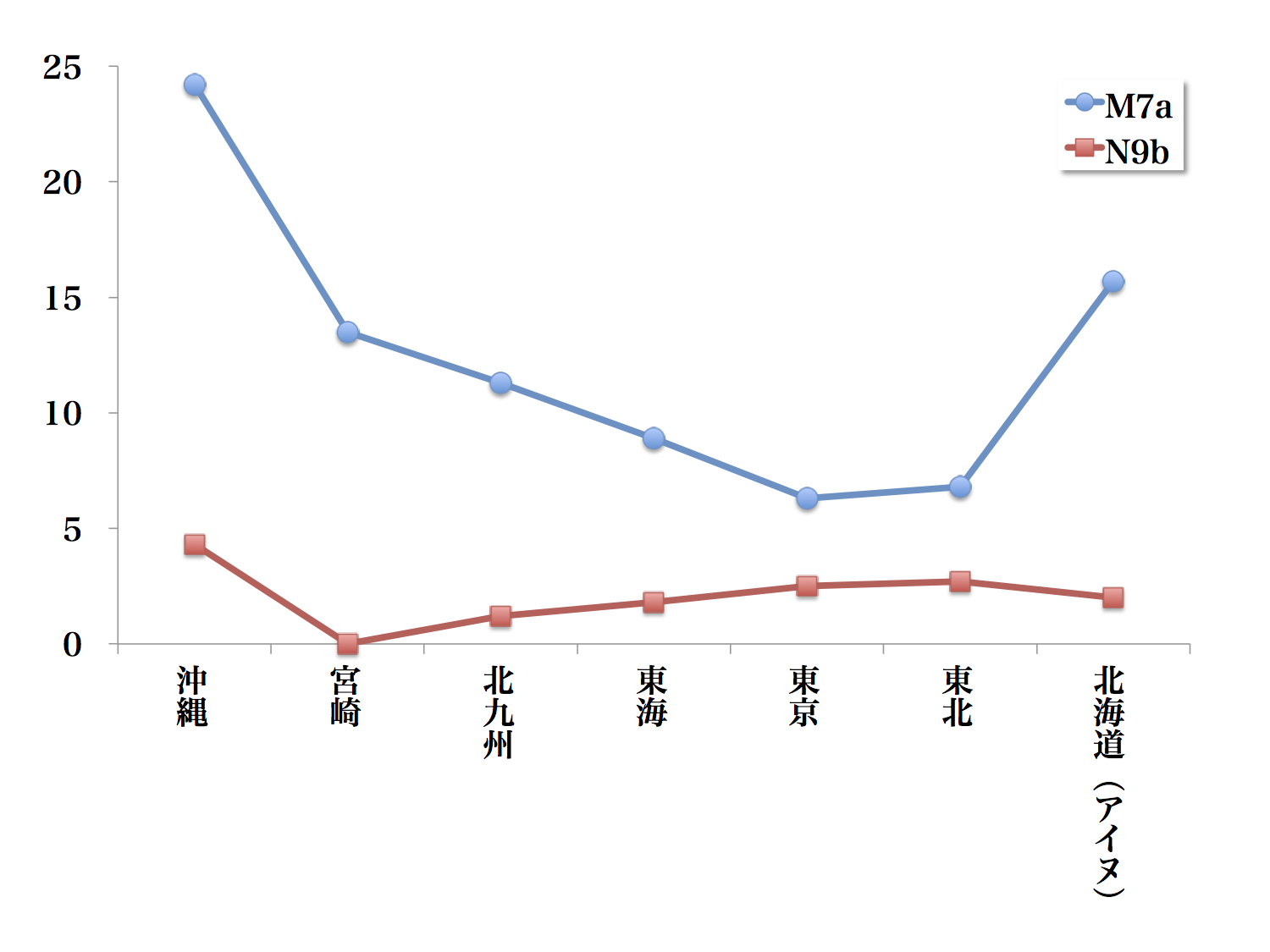
地域別に比較したM7aとN9bの頻度
篠田謙一著「日本人になった祖先たち」より改変
サブハプログループM7aは沖縄と北海道アイヌに多く日本本土の東京と東北でやや少ないM字型の分布傾向を示す。M7aは南から北に向かって減少する傾向を認める。ほかにN9b以外に、こうした一定の傾向を示すハプログループは存在しない。
サブハプログループN9bは沖縄に多く西日本に少なく東日本に多く分布している。南から北に行くほど徐々に頻度を増す地理的な勾配を認める。
篠田氏は「これが全国的な傾向として認められるのであれば、このハプログループは北から進入した基層集団によってもたらされたと考えられます」と述べている。しかしN9bは沖縄でも頻度が高い。はたしてその見解は正しいのだろうか。
古い基層集団であったとしても、九州から進入した新しい集団により徐々に南と北に移動を余儀なくされたと考えることも可能だろう。北に多いという傾向そのものは、N9bが北から進入したという根拠ではなく、基層集団として成立した後に起こった移動の結果であるとも考えられるのである。
ⅳ)東アジアにおけるミトコンドリアDNAハプログループの特徴
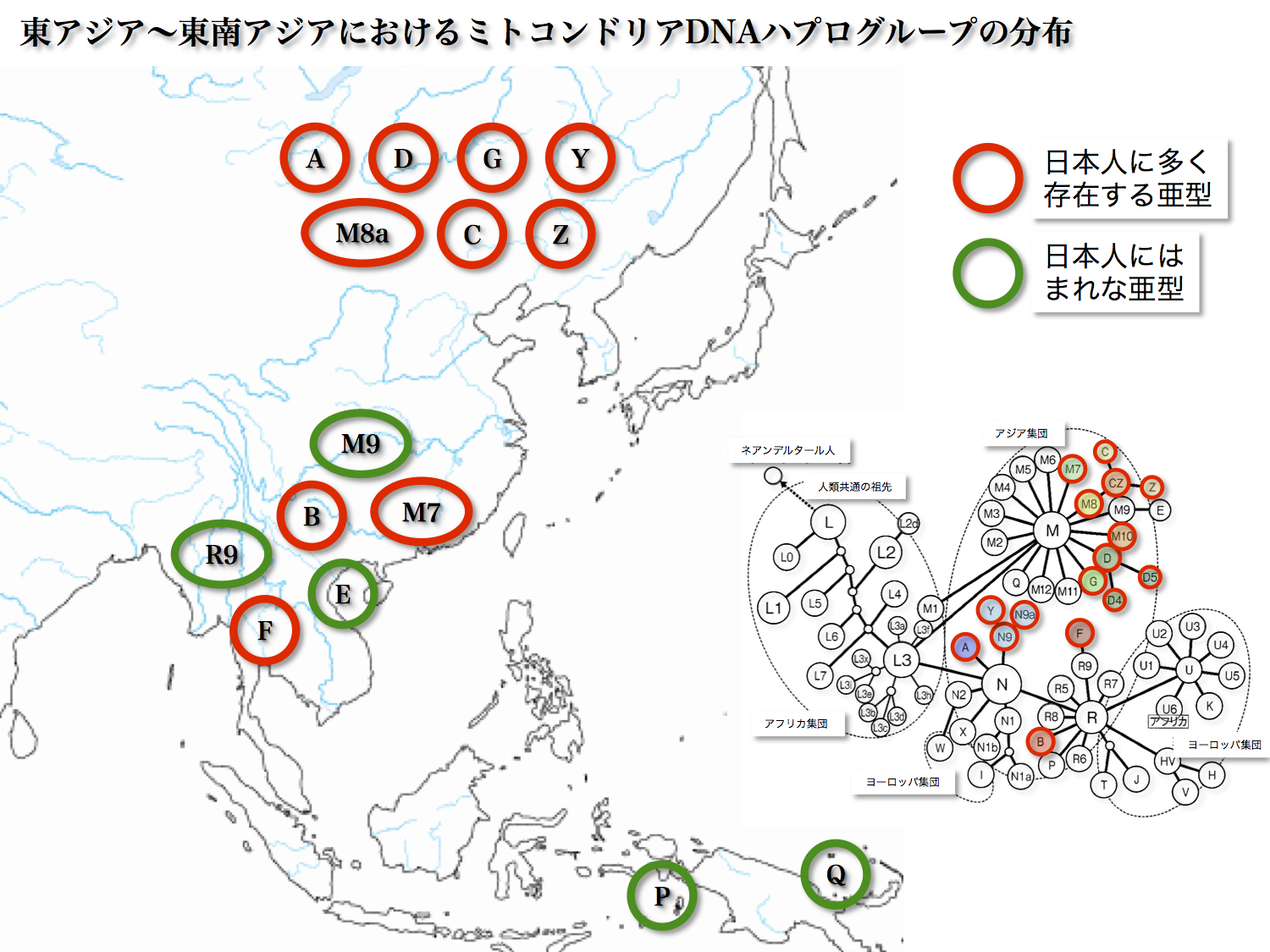
東アジア~東南アジアにおけるミトコンドリアDNAハプログループの分布
篠田謙一著「日本人になった祖先たち」より改変
東アジアから東南アジアにかけて分布している主なミトコンドリアDNAハプログループの分布を大まかに見ると図のようになる。東北アジアに主に分布するA、D、G、Y、M8aに対して、C、Z、M9、B、M7、R9、E、Fは主に東南アジアに分布する。
それぞれ北方系、南方系と呼び分ける人もいる。そのうち赤丸で示した集団が日本列島にも存在する。北方系のA、D、G、Y、M8a、C、Zと南方系のB、M7、Fである。
ちなみにオーストラリアにはP、ニューギニアにはQの分布が多い。
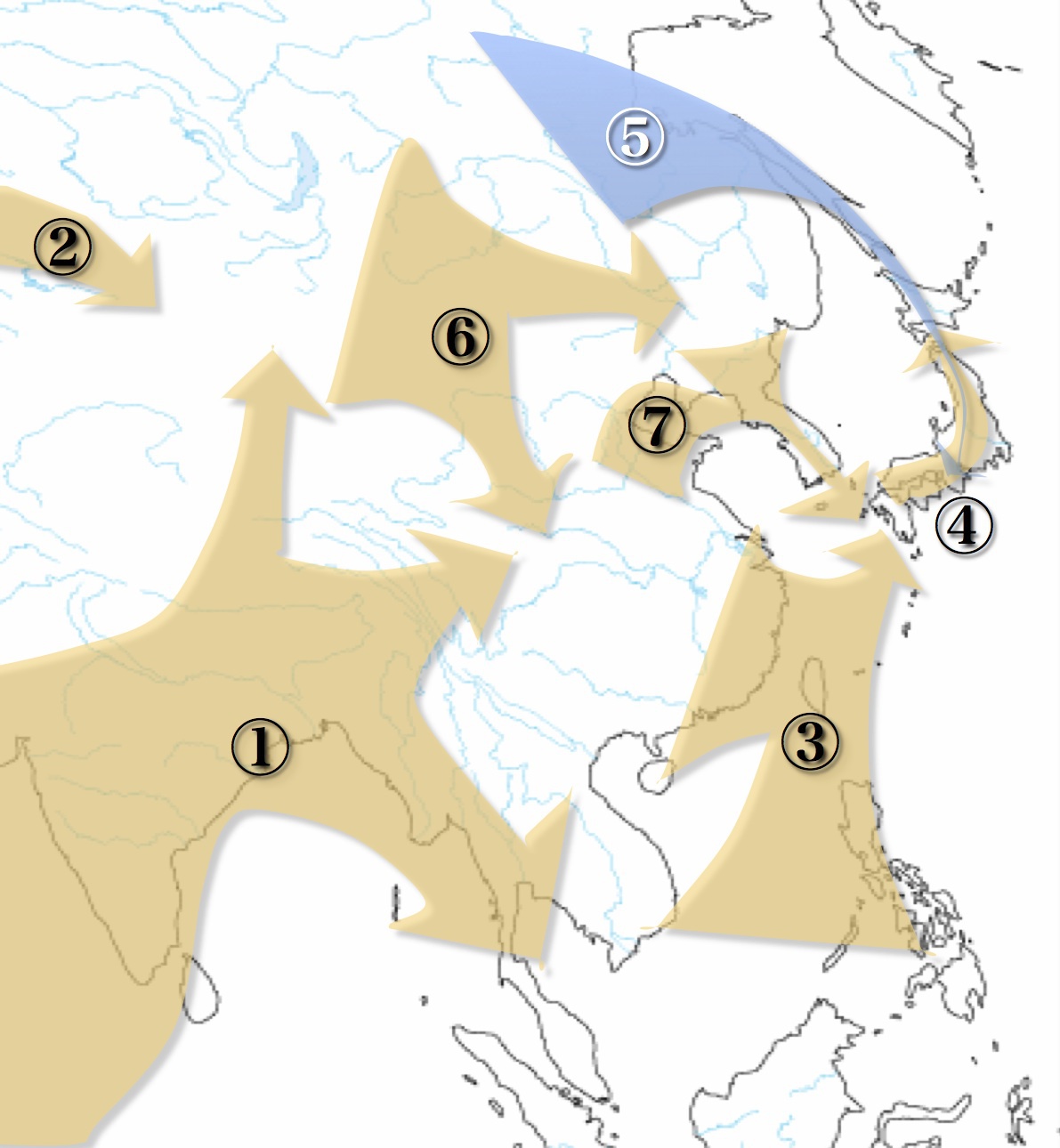
日本に移り住んだ祖先たちの経路想定図
ミトコンドリアDNAハプログループ研究によるもの
果たして真実はどうなのだろう?
後述のY染色体DNAハプログループ研究の成果と
合わせて再考されるべきだ
このように、北方系と南方系の人たちが、サハリン-北海道経由(図の⑤)、朝鮮半島経由(図の⑦)、東南アジア-華南から海を渡って(図の③)日本列島に移り住んだと考えるのが主流だった。果たしてその通りなのだろうか。Y染色体ハプログループ研究からは否定的である。次項とその次をご覧いただきたい。
 Soliloquy9
Soliloquy9