(11)日本人のルーツ Ⅱ Y染色体 OWLのひとりごと
![]()
日本人のルーツ(Ⅵ)Y染色体DNA研究のインパクト
2014.1.12
Y染色体DNA研究のインパクト 2014.1.12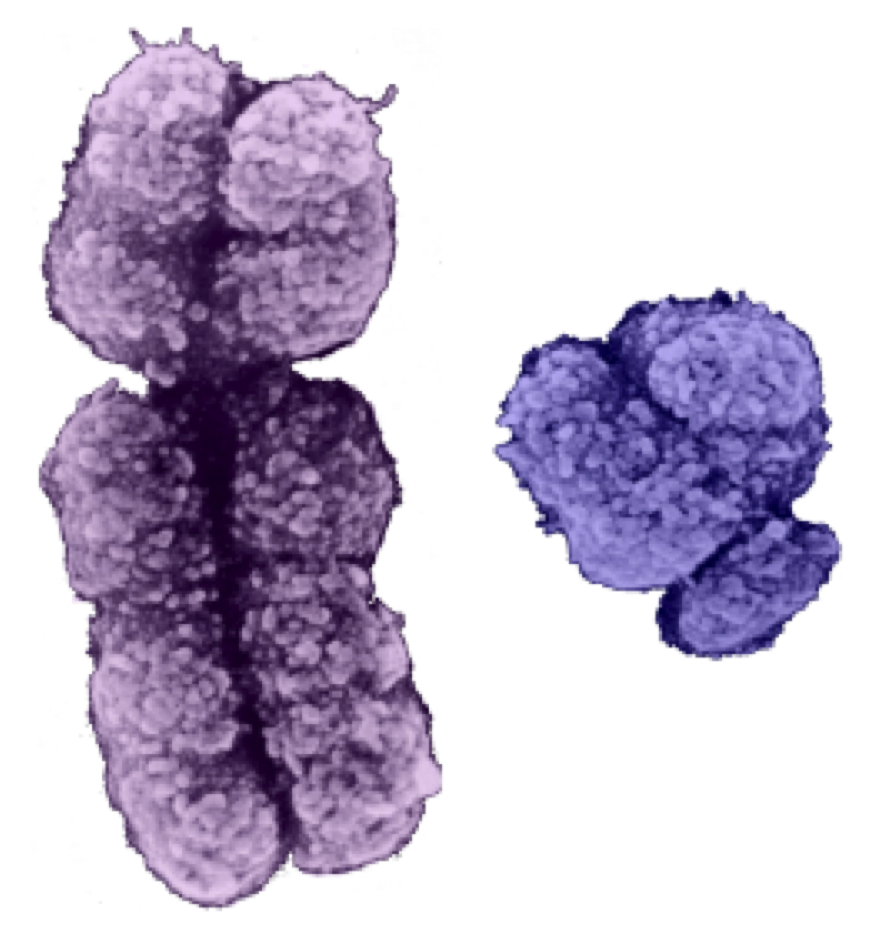
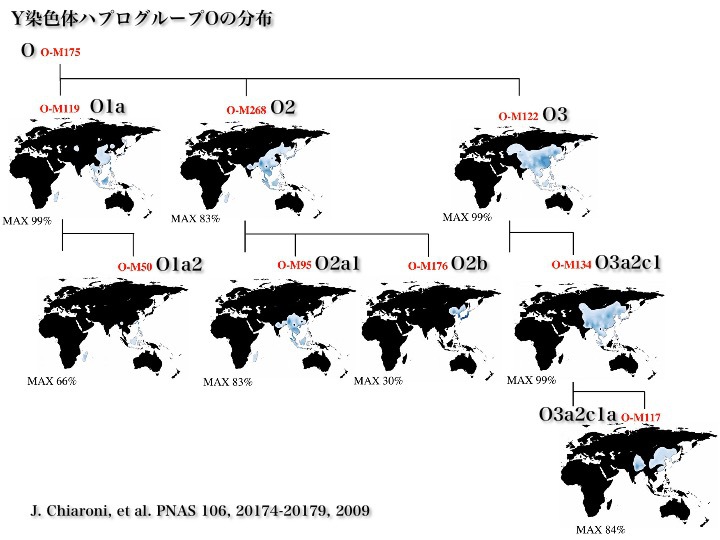
X染色体(左)、Y染色体(中)、Y染色体ハプログループ
本稿の文献はこちらを参照されたい。
Y染色体DNA研究のインパクト
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
このページのまとめ
1)Y染色体DNAにもハプログループが存在する。
2)Y染色体DNAハプログループの研究は、分子人類学に大きなインパクトを与えた。
3)ハプログループの系統樹が作成された。
4)単一起源説が支持された。
5)新人アダムの子孫は約7万年前に出アフリカを果たした。
6)特徴的なハプログループが全世界各地にわたって分布している。
7)出アフリカ後の人類の移動経路が推定された。
8)海岸線沿いに東へ向かったグループと
9)西アジアからヨーロッパに渡ったりインド亜大陸に向ったりしたグループがあった。
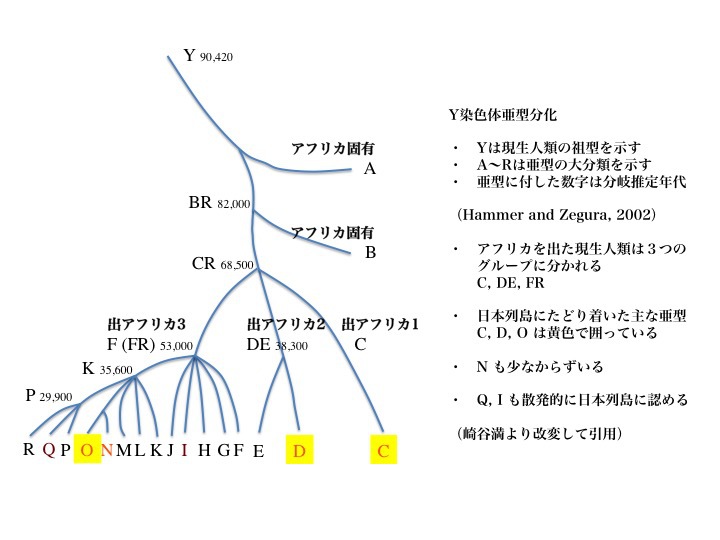
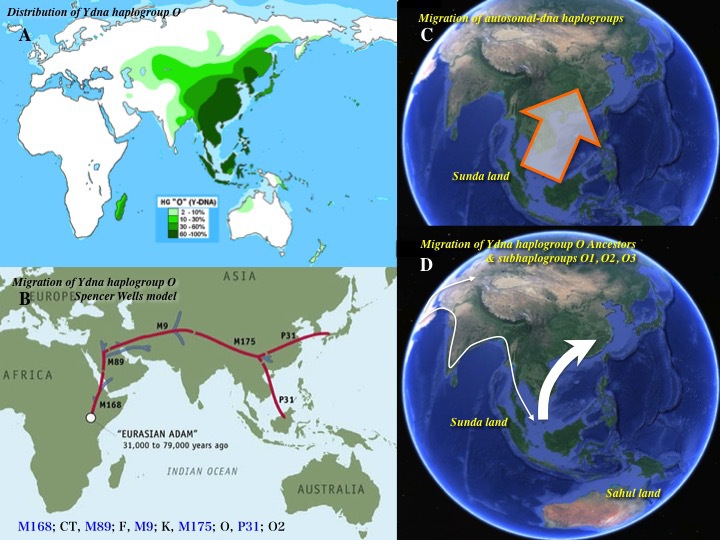
10)日本に来た主なハプログループは、C、D、Oである。
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
Ⅰ)Ydnaハプログループの登場と新人の単一起源説
ミトコンドリアDNAの研究に続いて、遺伝学に大きなインパクトを与えたのがY染色体の研究だ。その影響力は強く、ミトコンドリアDNA研究によって始まった分子遺伝学がよりいっそうの発展を遂げた。
Y染色体は父親から息子へと受け継がれる。ミトコンドリアDNAが母親がたの先祖を辿るのに対し、Y染色体は父親がたの先祖に迫ることができる。常染色体のように代を重ねるごとに薄まったりはしない。数万年という長い時代にわたって遡ることができる。
ただ今のところ、Y染色体の解析ができるのは生きたサンプルでなければならない。サイズの小さなミトコンドリアDNAの場合、考古学的な資料を使って解析可能なこともある。だがはるかに大きなサイズのYdnaの解析には、考古学的サンプルは使えない。
ミトコンドリアDNAと同様に、Ydnaにもランダムに点突然変異(mutation)が起こる。塩基がなくなったり(欠失=deletion)、塩基が入り込んだり(挿入=insertion)、他の塩基に変わったり(置換=substitution)する。300塩基にわたる挿入などの例(YAP=Y-chromosomal Alu Polymorphism)もある。
核のDNAはミトコンドリアDNAと比較して、突然変異を起こす確率は十分の一程度である。ミトコンドリアDNAの方が変異の速度が早い。核のDNAは変異の速度が遅く、長い過去にまで遡ることが容易だと言われる。
ただ、そもそもYdnaの長さはミトコンドリアDNAに比べて3000倍ほどある。結果的に、Ydnaには多量の突然変異が蓄積されていると考えて良い。
父親に二人の息子がいるとして、兄に受け継がれたY染色体のどこかに変異が起こり、弟には起きなかったとする。兄の子孫には代々その変異が受け継がれ、弟の子孫は父と全く同じタイプが受け継がれる。
代を重ねると他の部位でも変異が起こり、そのまた子孫に受け継がれる。こうして変異自体が蓄積されてゆくことになる。上述の祖先の弟にも変異が起きたとしても、兄に起きた最初の変異と全く同じ部位で同じ塩基に置き換わったり、全く同じ塩基が欠失したりする確率はほぼゼロである。
代々積み重なった変異の組み合わせパターンを解析することによって、Ydnaはさまざまなグループに分けられる。そのパターンはミトコンドリアDNAの場合と同様に、ハプログループと呼ばれる。
Ydnaハプログループの解析により、共通祖先(MRCA)の系統樹が作成された。ミトコンドリアDNAの系統樹と同様である。次の図は、左がY染色体ハプログループ、右がミトコンドリアDNAハプログループの系統樹を示している。
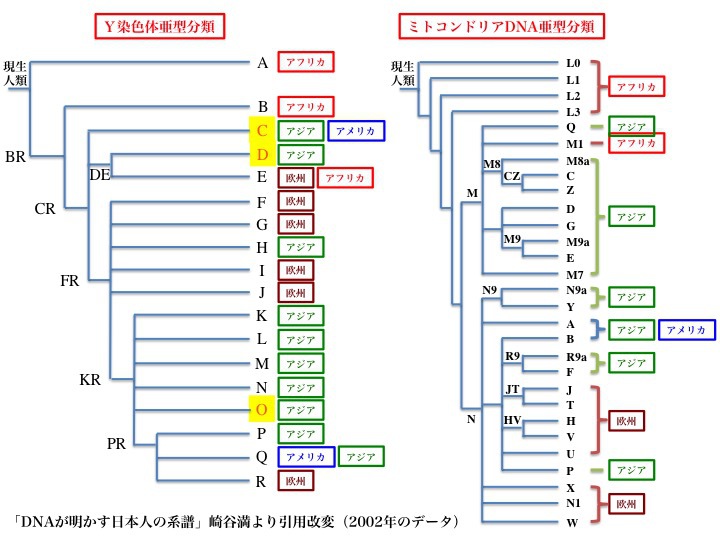
Ydnaハプログループ(左図)とミトコンドリアDNAハプログループ(右図)それぞれの系統樹の比較(2002年のデータ)
2002年のデータによると、Y染色体ハプログループはAからRまで18亜型に分類される。図で、Aはアフリカ、Cはアジアとアメリカ大陸など、分布している主な地域が記されている。
Ydnaのハプログループ解析により、Y染色体の共通祖先は約九万年(プラスマイナス二万年)前に誕生したと言われる。出アフリカを果たしたのは68000年前と推定されている。これらはミトコンドリアを用いたデータと基本的に一致している。
ミトコンドリアDNAの研究と同様、15~16世紀に始まった植民地時代より以前の分布で表している。アメリカ、オセアニアなどはそれぞれの先住民のことである。
左と右の系統樹を比較すると、アフリカにとどまったグループ、アジアに移動したグループ、アジアとアメリカに展開したグループ、ヨーロッパに行ったグループが存在している。違うサンプルを異なった方法で解析したのに、順番に多少の相違はあるものの、驚くほどよく対応していることがわかる。
祖先から分岐し、それからまた別のハプログループに分岐してゆく際に、たとえばDとEが分岐する前の親ハプログループをDE、FからRまで分岐する親ハプログループをFRなどと呼ぶ。他にKR、PRも最終分岐前の親ハプログループになっていることがわかる。
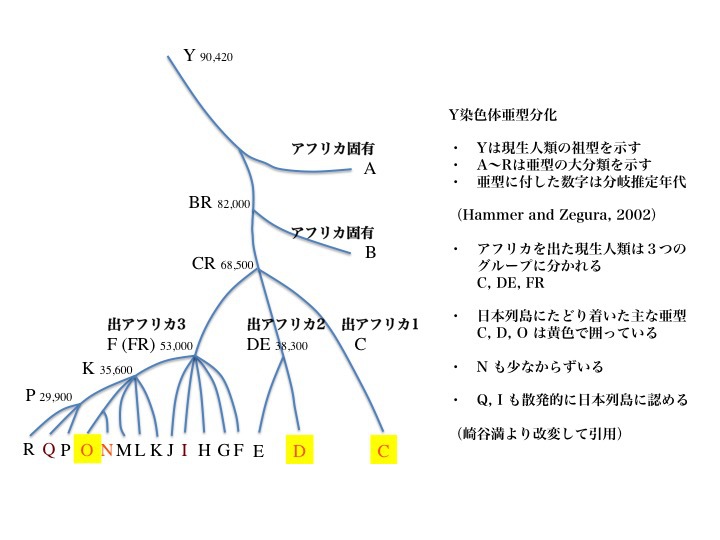
アダムの子孫の出アフリカと日本にたどり着いた主なハプログループ
大規模な解析から、Y染色体は一人のアダムに遡ることができることが判明した。ミトコンドリアDNAが一人のイブにたどり着くのと同様である。アフリカで多く分岐したあと、三つのグループが出アフリカを果たしたと想定されている。ハプログループCとDEとFR。年代は約6万年前と推定された。
日本列島に辿り着いたハプログループは、C、D、Oが主な亜型で、黄色で囲ってある。N、Q、I なども散発的(sporadic)に日本列島に認める。
Ⅱ)Ydnaハプログループ研究の進展と世界分布
研究が進むにつれて、Ydnaハプログループのルートに関する仮説は修正されていった。
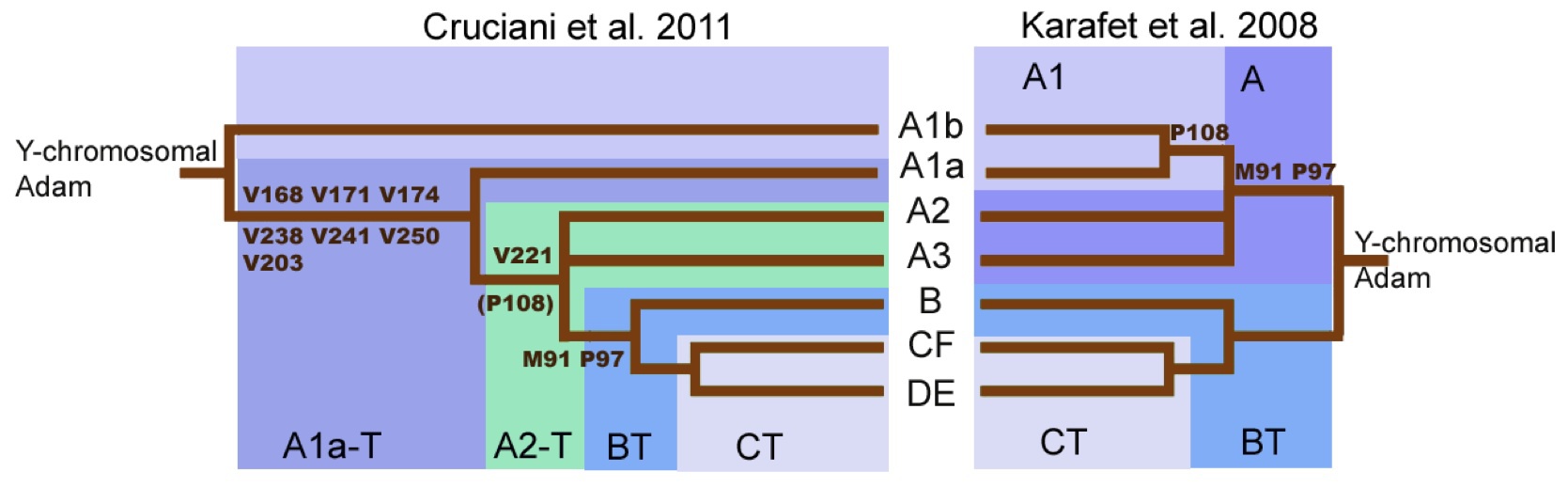
クルシアーニらによるYdnaのルートに関する仮説(図左側:Cruciani 2011)とカラフェットらによる説(図右側:Karafet 2008)の比較 Wikipediaによる(CC BY-SA 3.0)
Y染色体アダムに関する系統樹は、かつて図の右のように考えられていた。2011年には図の左のような仮説が提唱された。左側の説はメンデスら(Mendez 2013)の研究によって検証され、さらに2013年の国際遺伝系図学会(ISOGG=The International Society of Genetic Genealogy)でさらに修正された。
Y染色体アダムばかりでなく他のハプログループの研究も進み、2013年時点での系統樹は次の通りとなった。
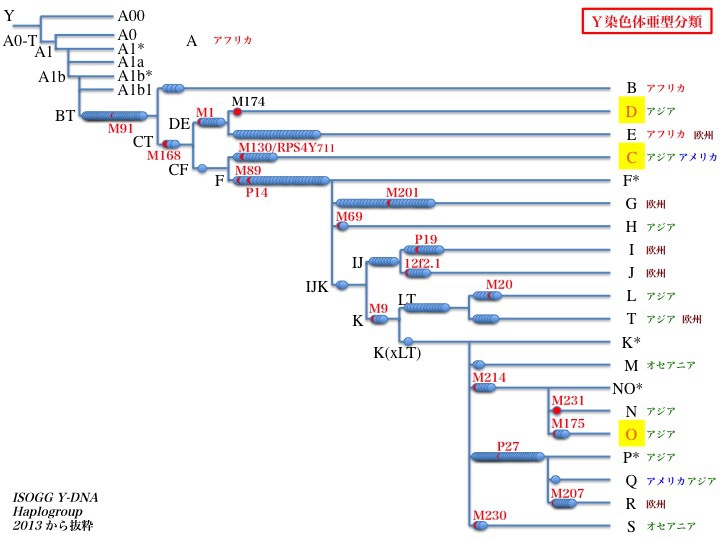
2013年のデータによるYdnaハプログループとその世界分布
現在確認されている変異一個をブルーの点一つで表している。赤の点とM91やM174などは、分類に頻用されている有名な変異の名称である。
2002年のデータに比べて次の点が変更になっている。
1)SとTが加わっている。
2)Fを親としてG、Hの他に IJK が連なっている。
3)IJKから IJとKに分岐している。
4)KはLTとK(xLT)(LTが抜けた残りのK)に続く。K(xLT)はMNOPSとも呼ばれている。
5)LTからLとTが分かれている。
6)K(xLT)を親としてM、NO、PQR、Sが分岐している。
ハプログループAとBをのぞいた残りのグループCTの変異は、アフリカ北東部のどこかで起こった。そのあとCTの一部が出アフリカを果たす。
ハプログループDEに見られる変異とCFに見られる変異は、65000年前頃に起こったと推定される。DEの変異はアフリカで起こり、Eの一部とDの集団が出アフリカを果たしたのだろう。CFの変異はアジアで発生し、その集団が世界の大部分の人類の父祖となった。
各ハプログループの推定分岐年代、現分布地域がこちらのテーブル「Y染色体DNA各ハプログループの推定発生年代と場所」にまとめてある。参照されたい。
ハプログループはさらに多くのサブハプログループに別れ、D2、O2bなどと呼称されている。2013年現在の詳細系統樹はここを参照されたい。
ⅰ)世界各地域におけるYdnaハプログループ構成頻度
出アフリカを果たした人々が世界各地に移動拡散していった足取りを見てみよう。植民地時代が出現する以前、先住民はどのように分布していたのだろうか?その様子が、現代におけるY染色体ハプログループの分布から推定されている。
まずは全世界におけるY染色体ハプログループの分布頻度を概観する作業から始める。
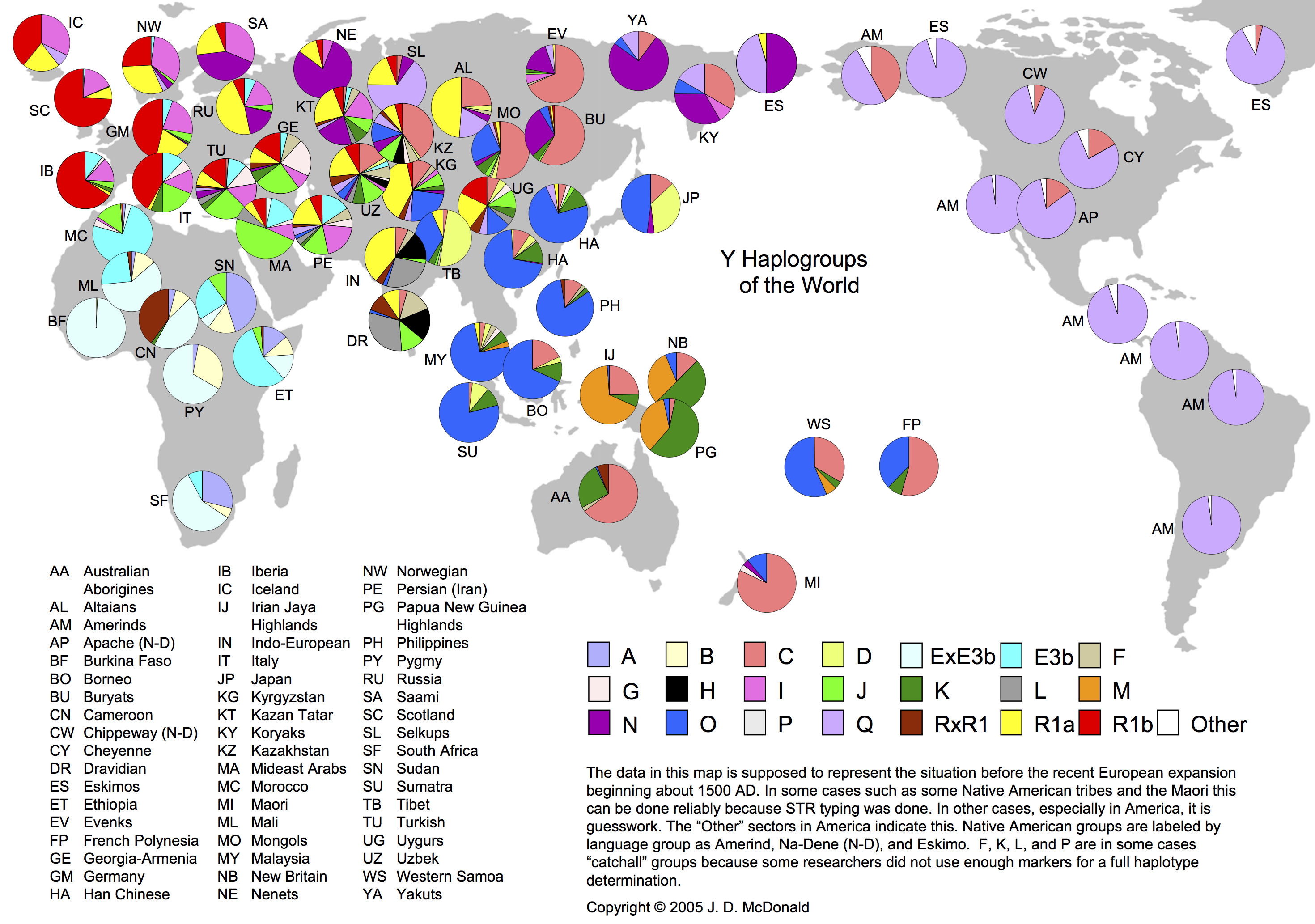
Y染色体ハプログループの世界分布
http://www.scs.illinois.edu/~mcdonald/WorldHaplogroupsMaps.pdf より
さらに大きな図はこちら
(a)アフリカ大陸
ハプログループE(ライトブルーとうす青色)が圧倒的に多い。例外はスーダンのA(藤色)である。カメルーンのR(茶色)はEと同程度分布している。ピグミーにはEに続いてB(うす黄色)が多い。モロッコにはヨーロッパに見られるJ(緑)も一部存在する。
(b)ヨーロッパ
サブハプログループR1(赤、黄色)がヨーロッパを特徴づけている。西がR1b(赤)優位で、東(ロシア)がR1a(黄色)優位、北が両者拮抗という傾向。イタリアではR1b優位なところに I(ピンク)やJ(緑)が混じる。スカンジナビアでは I が多く、アイスランド、スコットランド、ロシアにも一定割り合いの I が存在する。
スカンジナビア北部のサーミ(Saami)やロシア極北部のネネッツ(Nenets)にはN(赤紫)が多数派を占める。
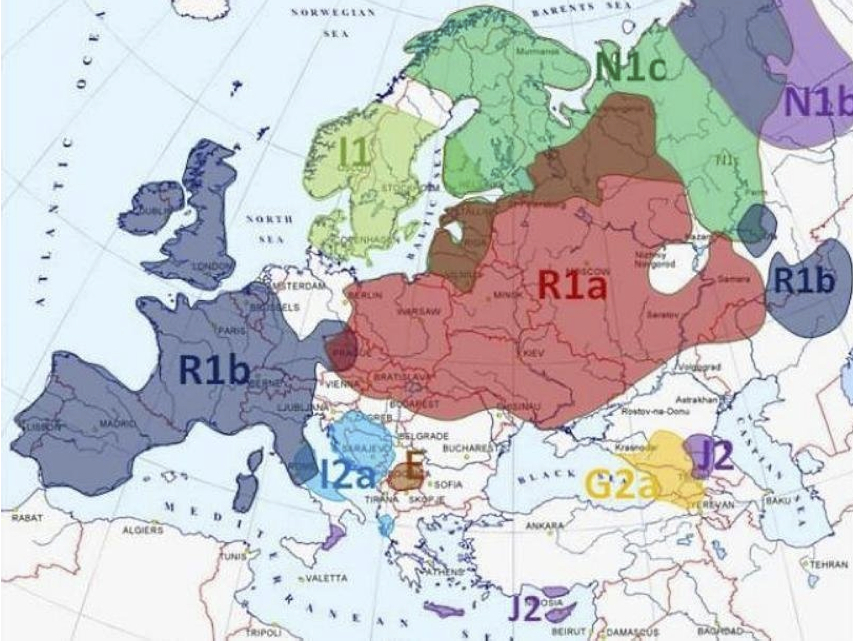
ヨーロッパでの主要なYdnaハプログループの分布傾向
http://www.geocurrents.info/category/place/europe/page/2 による。
この図は、ヨーロッパ各国の主要Ydnaハプログループの地域分布傾向をわかりやすく示している。英、仏、ベルギー、南ドイツ、スイス、北部~中部イタリア、イベリア半島でR1bが最も多く、ポーランド、チェコ、スロバキア、ベラルーシ、ウクライナ、ロシア西部でR1aの頻度が最も高い。
アドリア海をはさんで中風イタリアとクロアチア、セルビア、ボスニア・ヘルツェゴビナには I2aが分布する。コソボがE主体であることは驚きだ。
ノルウェー、スエーデン、デンマーク三国は I1、フィンランドとスカンジナビア半島北部にN1cの集団が分布する。
他にコーカサス地方では、グルジアがG2a、チェチェンがJ2であること、地中海ではイタリア半島突端のカラブリア州、ギリシャのクレタ島、トルコの南キプロス島の住民多数派がJ2であることも興味深い。
再びもとの円グラフ(パイチャート)が沢山載っている世界地図に戻って説明する。
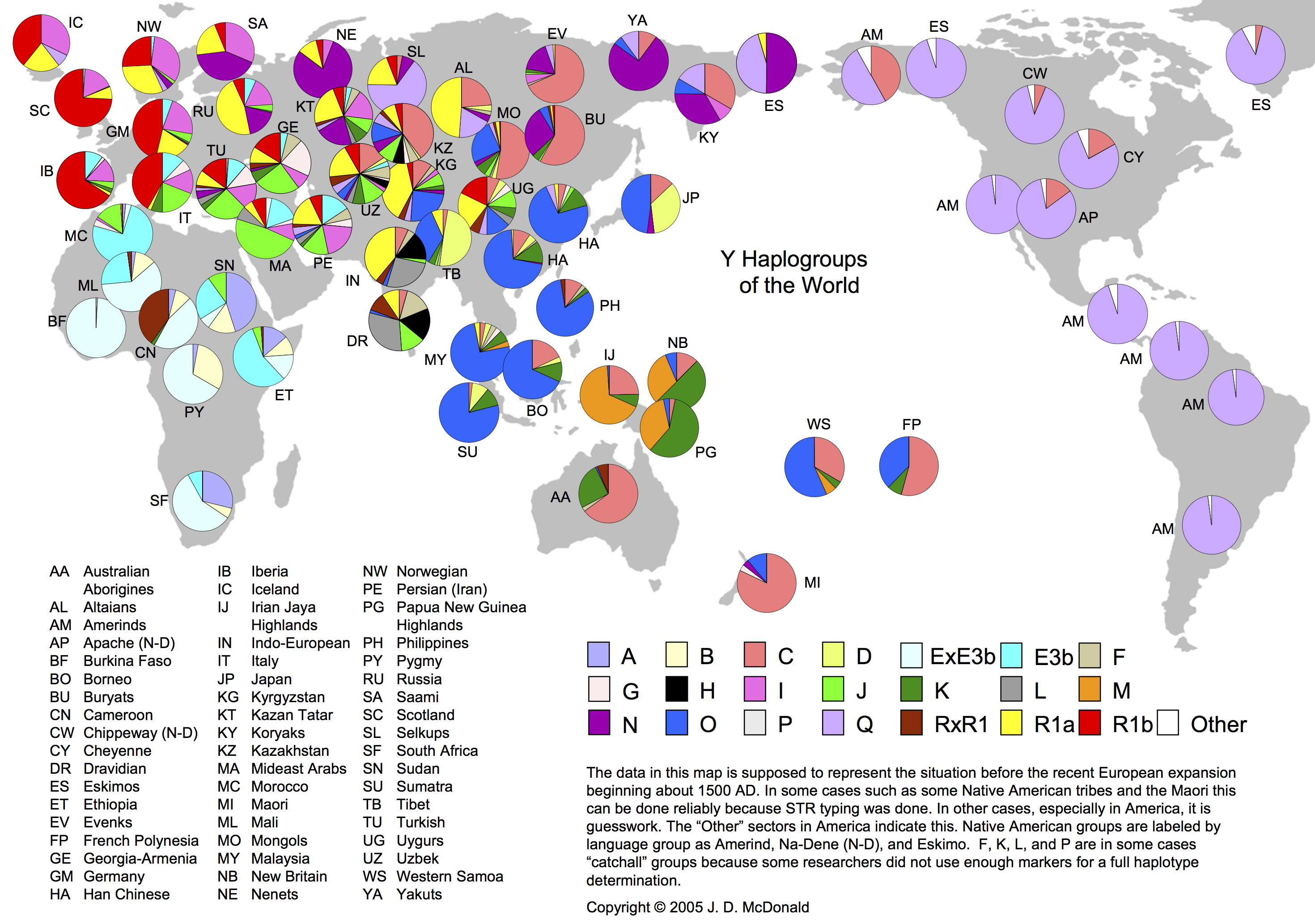
さらに大きな図はこちら
(c)西アジア
トルコ、アルメニアではJ(緑)とR1(赤、黄色)が多い。次いで多いのがトルコでは I(ピンク)、G(淡い桜色)の順、アルメニアでは逆にG、I の順。
中東のアラブ人にはJ(緑)が多く、次いでE(ライトブルー)、I の順。
ペルシャ人は I 、R1の頻度が高く、次いでEとJで、四つで人口の大部分を占める。
西アジアの人々は総じてR1、J、I 、E、Gが混じった集団と言える。
(d)南アジア
北インドではR1a(黄色)、L(灰色)、H(黒)の順、南インドのドラビダ人はL、H、R(茶色と黄色)、F(ライトブラウン)、J(緑)の順でそれぞれ大部分を占める。どちらにも少数ながらCが混じっている。
(e)東南アジア、華南、華北
圧倒的にO(青)が多い。K(ダークグリーン)が続く地域が多く、スマトラではD(うす黄色)、ボルネオやフィリピンや華南ではC(赤みがかったオレンジ色)の頻度も目立つ。
(f)日本
ハプログループO(青)、D(うす黄色)の順で八割ほどを占め、C(赤みがかったオレンジ色)がそれに続く比較的単純な構成。他の東アジアや東南アジアとは全く様相が異なる。
(g)チベット
ハプログループD(うす黄色)、O(青)の順で日本の逆となり、全体の九割ほどを占める。華南、華北と全く違う民族と言って構わない。
(h)ウイグルと中央アジア
シルクロード地方だけあって人種の坩堝といった様相。ウイグル、キルギスタン、ウズベキスタンで最も高頻度なのはR1(黄色と赤)。カザフスタンで最も多いのがC(赤みがかったオレンジ色)。東アジアに近いほどO(青)が、モンゴルや北アジアに近いほどCの割合が多い。J(緑)も一定割合存在する。少数のハプログループが他にもたくさん存在する。
(i)北東アジア
モンゴル、エヴェンキ、ブリヤートではC(赤みがかったオレンジ色)が多数派。モンゴルではO(青)が次ぎ、エヴェンキとブリヤートではN(赤紫)が続く。アルタイではR1a(黄色)が半数を占め、C、Q(うす紫)の順で大多数となる。
極北ではNが多く、ヤクートでは七割以上、エスキモーで半数、コリヤークで三割を占める。Nと匹敵するほど多いのがエスキモーでQ、コリヤークではCとなる。
(j)アメリカ大陸
アメリカ大陸先住民の多くを占めるのがQ(うす紫)である。アラスカのアメリンドで約半数、シャイアンで七割強、アパッチで八割近く、チペワやエスキモーで九割を占めるばかりか、それ以外ではほぼ100%がQである。
第二グループでは、アメリンドの四割、シャイアンやアパッチの二割前後がC(赤みがかったオレンジ色)である。ミトコンドリアDNAハプログループの研究から、アメリカ大陸の女性は多様性に富むことが判明している。女性と違って男性は、極端な淘汰がなされたと考えざるを得ない。
(k)オセアニア
各地域で差が大きい。オーストラリアではC(赤みがかったオレンジ色)が六割以上、K(ダークグリーン)、R(茶色)が続く。
ニューギニアを含むメラネシアでも地域差が激しい。パプアニューギニアやニューブリテン島ではKが多数で、M(オレンジ色)が続いて八~九割を占める。ニューギニア島の西側イリアンジャヤではMが七割でCが次に多く、二つで全体の九割以上となっている。
ニュージーランドを含むポリネシアでも地域差が大きい。ニュージーランドではCが八割近くを占め、仏領ポリネシアではCが半数以上。次いでO(青)が多い。西サモアではOが半数以上と多数派で、Cが三割と続く。
ⅱ)各ハプログループの世界分布
次いで、各ハプログループごとに世界のどの地域に分布しているか、一つひとつ検討してゆこう。分布を調べた上でそれぞれがどういう移動経路をたどったのか推定してみる。
(a)ハプログループDとE
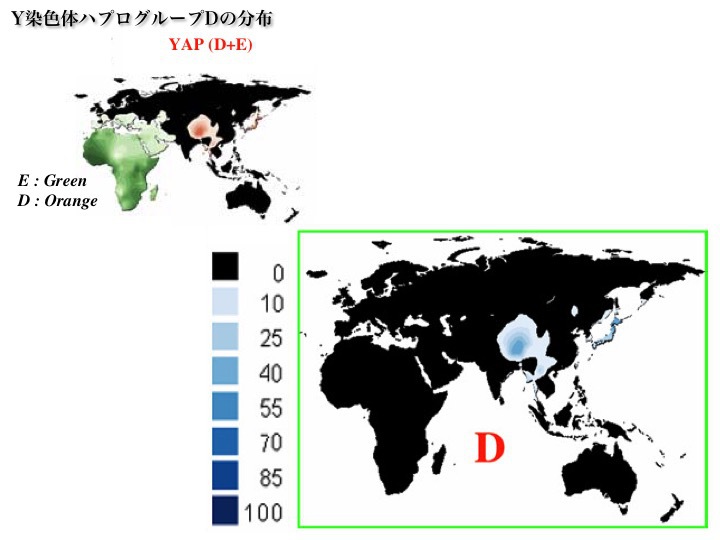
ハプログループDの分布
Chiaroni 2009より改変
ハプログループDは50000年前に発生。日本、チベットに多く特徴的。他にインド洋のアンダマン諸島、東南アジア、モンゴル、中央アジア、南シベリアなどにも少数分布している。
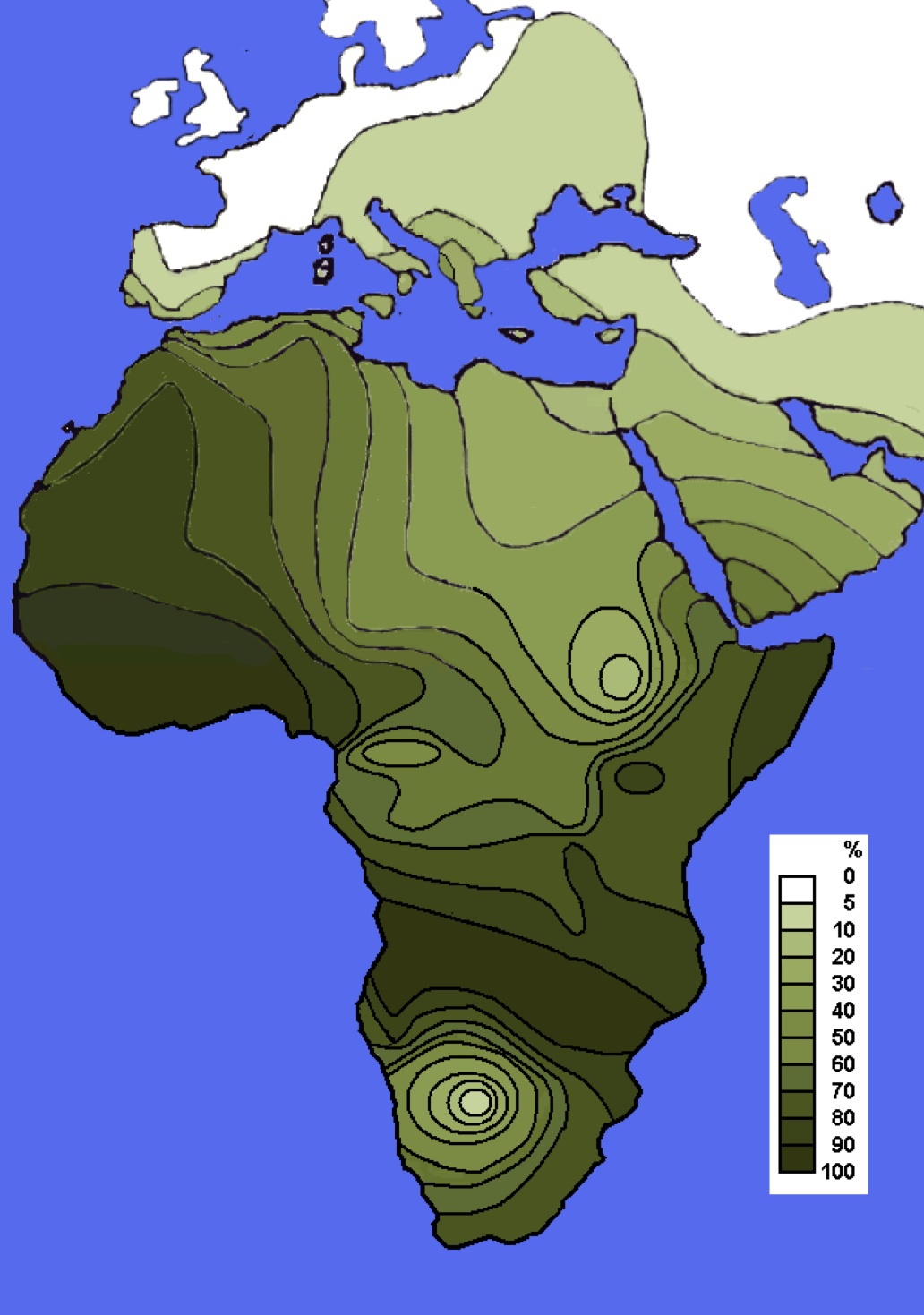
ハプログループEの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループAとBとならんで、Eはアフリカ大陸に多い。西アジア、南ヨーロッパ、東ヨーロパにも見られる。
(b)ハプログループC
ハプログループCには次の七つのサブハプログループが存在する。
C*、C1、C2、C3、C4、C5、C6。
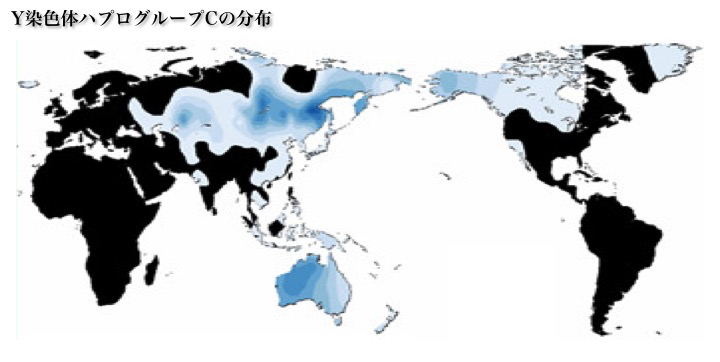
ハプログループCの分布
Chiaroni 2009より改変
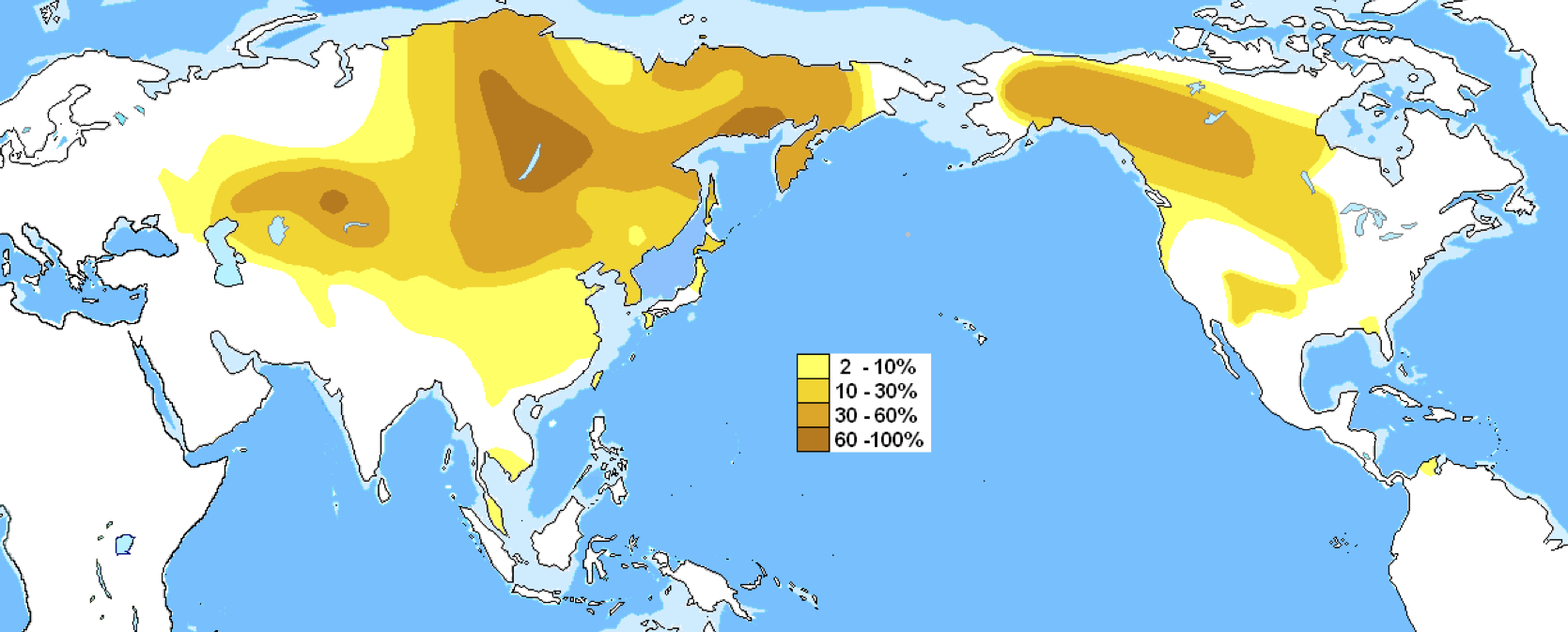
ハプログループC3の分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループCのうちサブハプログループC3は、シベリアなど北東アジア、中央アジア、北米に拡がっている。C3の他、インドにそのままとどまったC*、C5、オーストラリア大陸に行ったC4、ニューギニアに向かったC2、C5、オセアニアに達したC*、C6、日本に行きそこにとどまったC1サブハプログループがある。現在の分布から特徴的な移動を追うことができる。
(c)ハプログループG
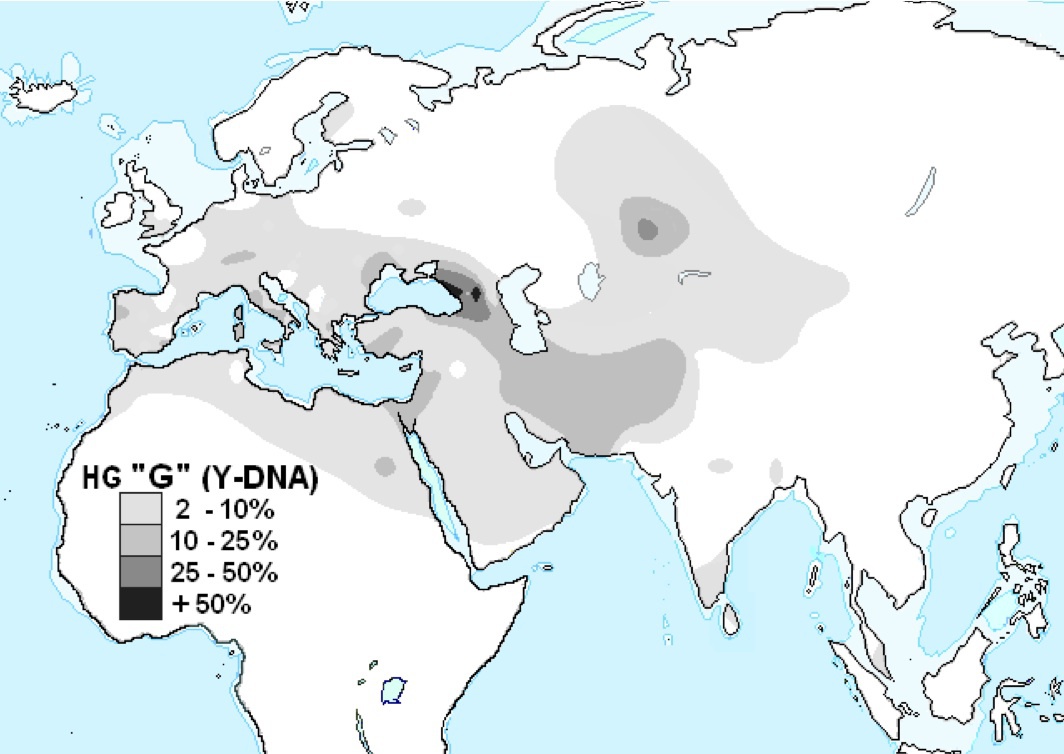
ハプログループGの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループGはコーカサス地方に多く、西アジアから中央アジアに拡がる一方、西は地中海を囲むようにしてヨーローッパに拡がっている。
(d)ハプログループH
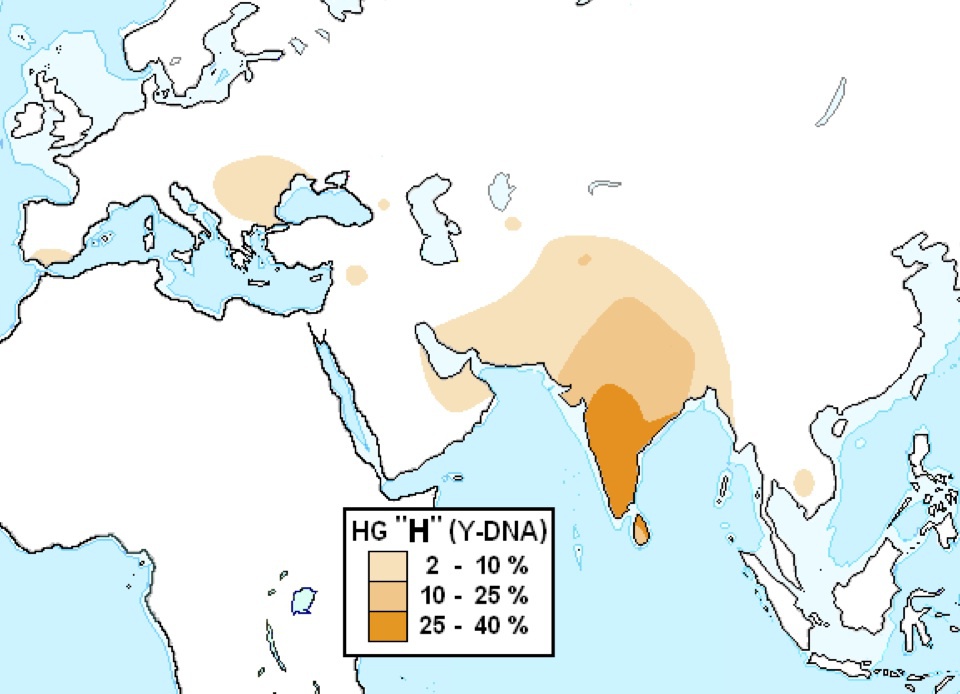
ハプログループHの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループHは南アジア特にインド南部を中心に分布している。
(e)ハプログループ I とJ
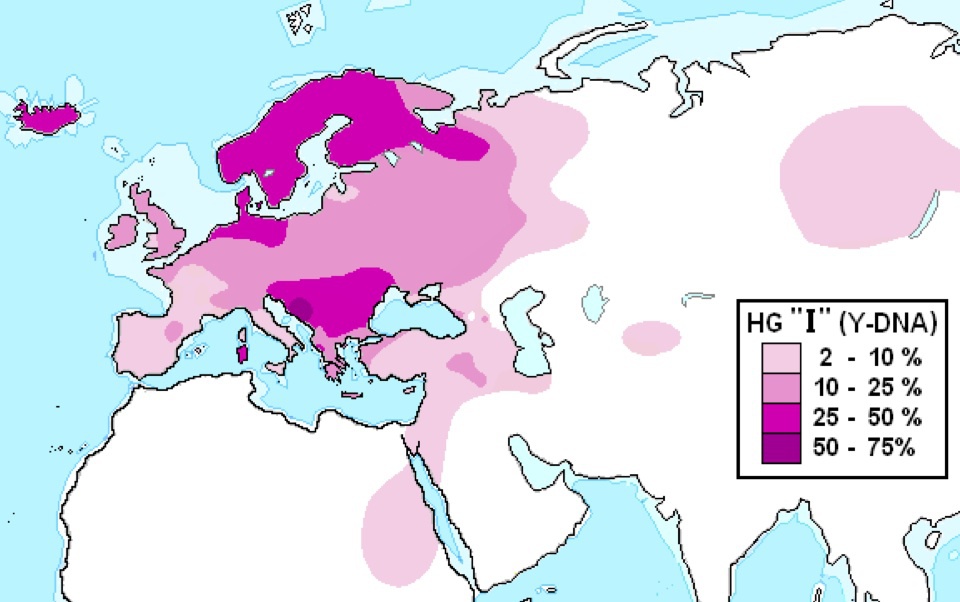
ハプログループ I の分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループ I はバルカン半島、デンマークから北ドイツ、スカンジナビア三国、アイスランドに多く、ヨーロッパ各地域ならびにトルコなど西アジア、シベリアにも分布する。
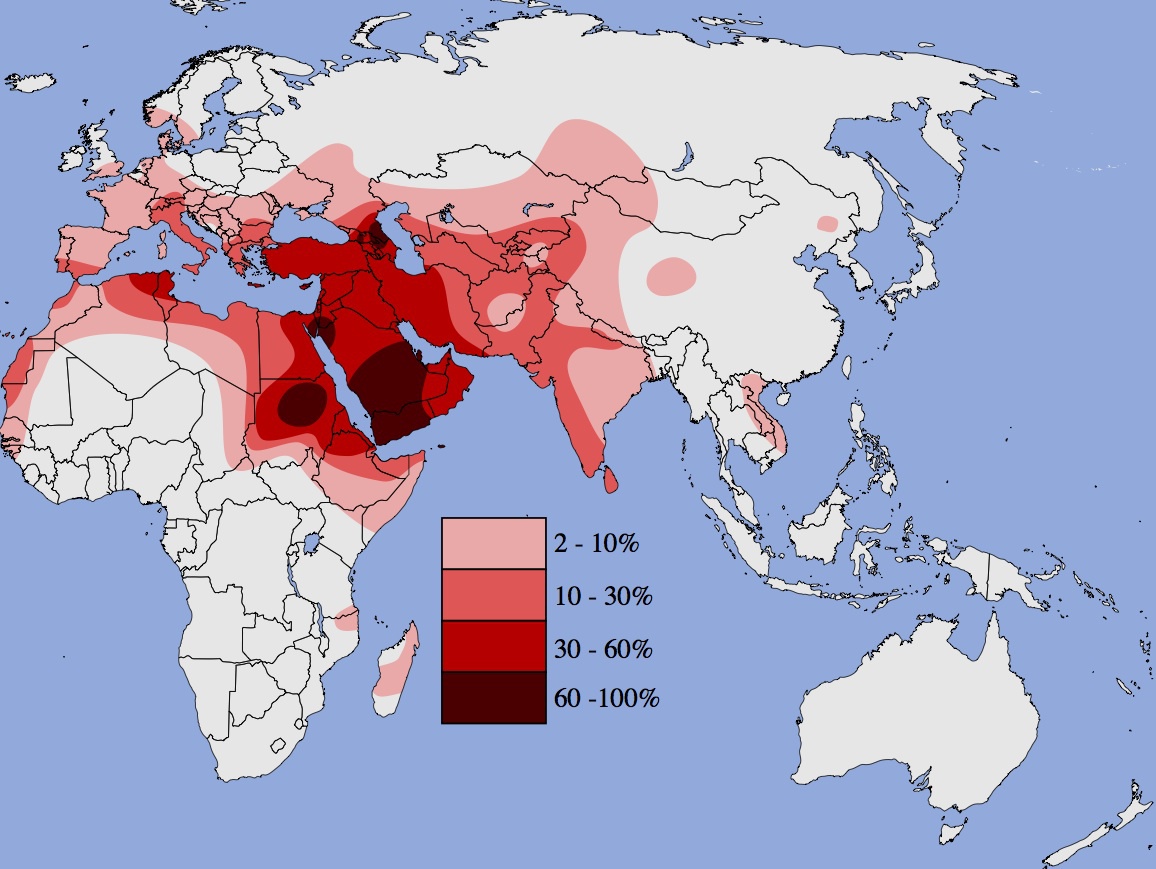
ハプログループJの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループJは西アジアを中心に、地中海沿岸、黒海周辺、中央アジア、南アジアに拡がっている。
(f)ハプログループLとT
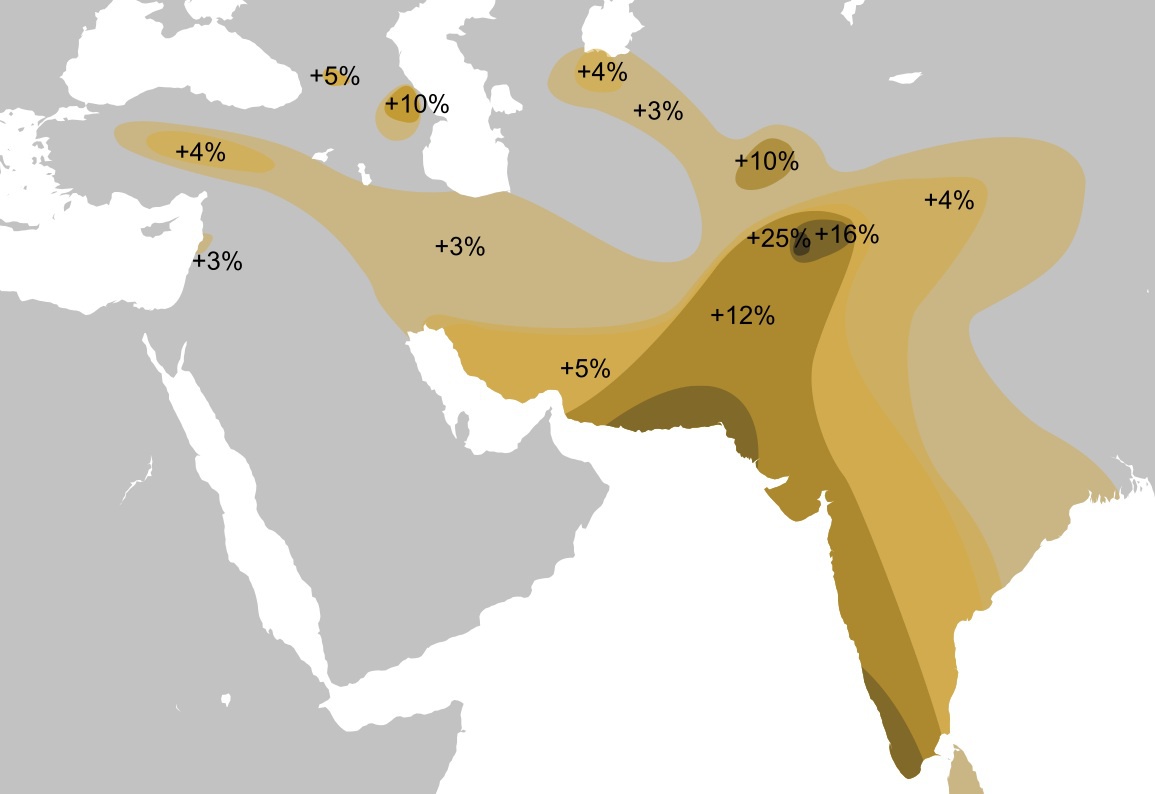
ハプログループLの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループLはパキスタン地方インダス河沿いを中心に、その東西に分布が広がっている。
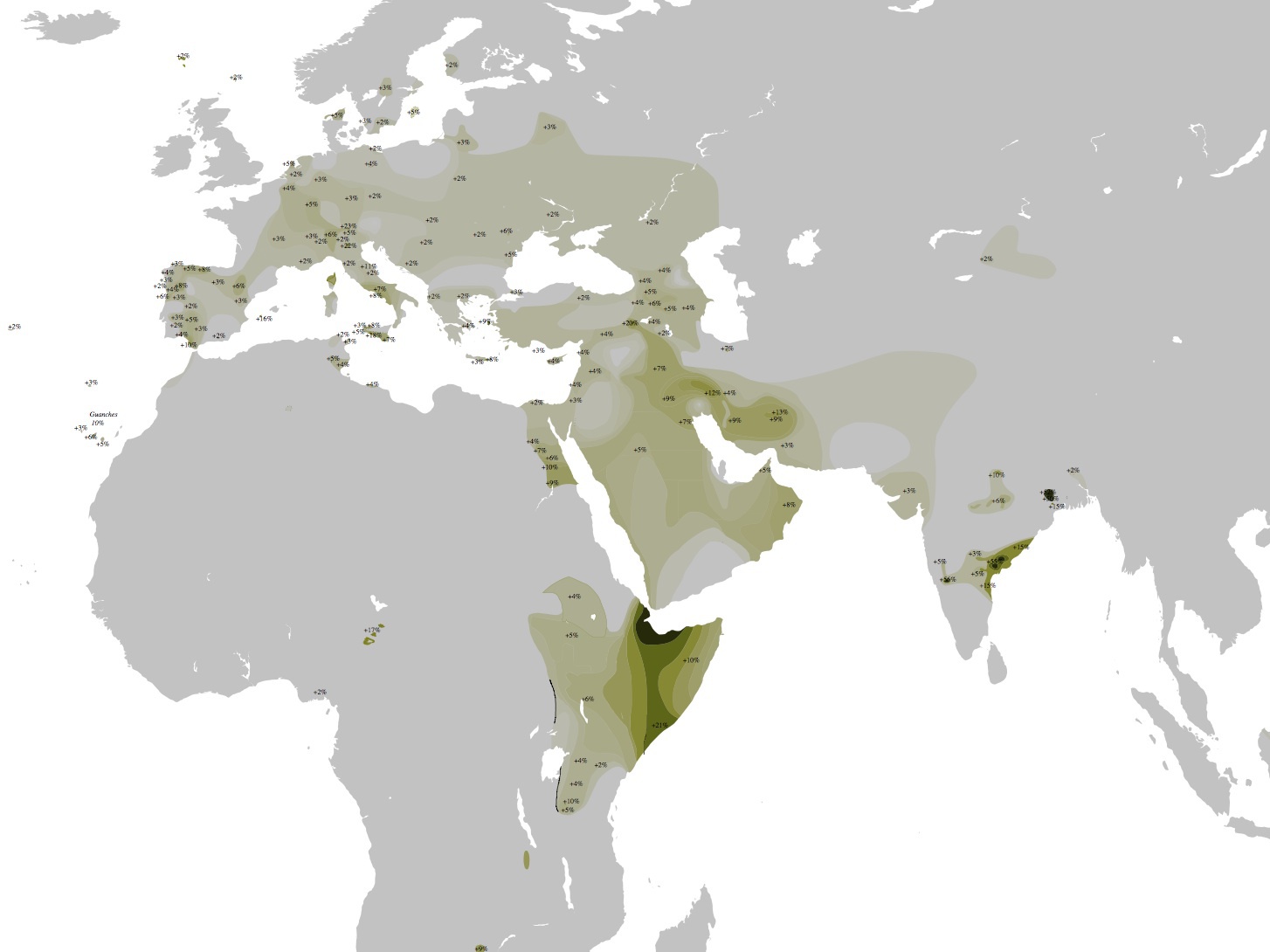
ハプログループTの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループTはエチオピア、イラクやイラン南部を中心とした西アジア、ヨーロッパ南部から東部に見受けられる。
(g)ハプログループNとO
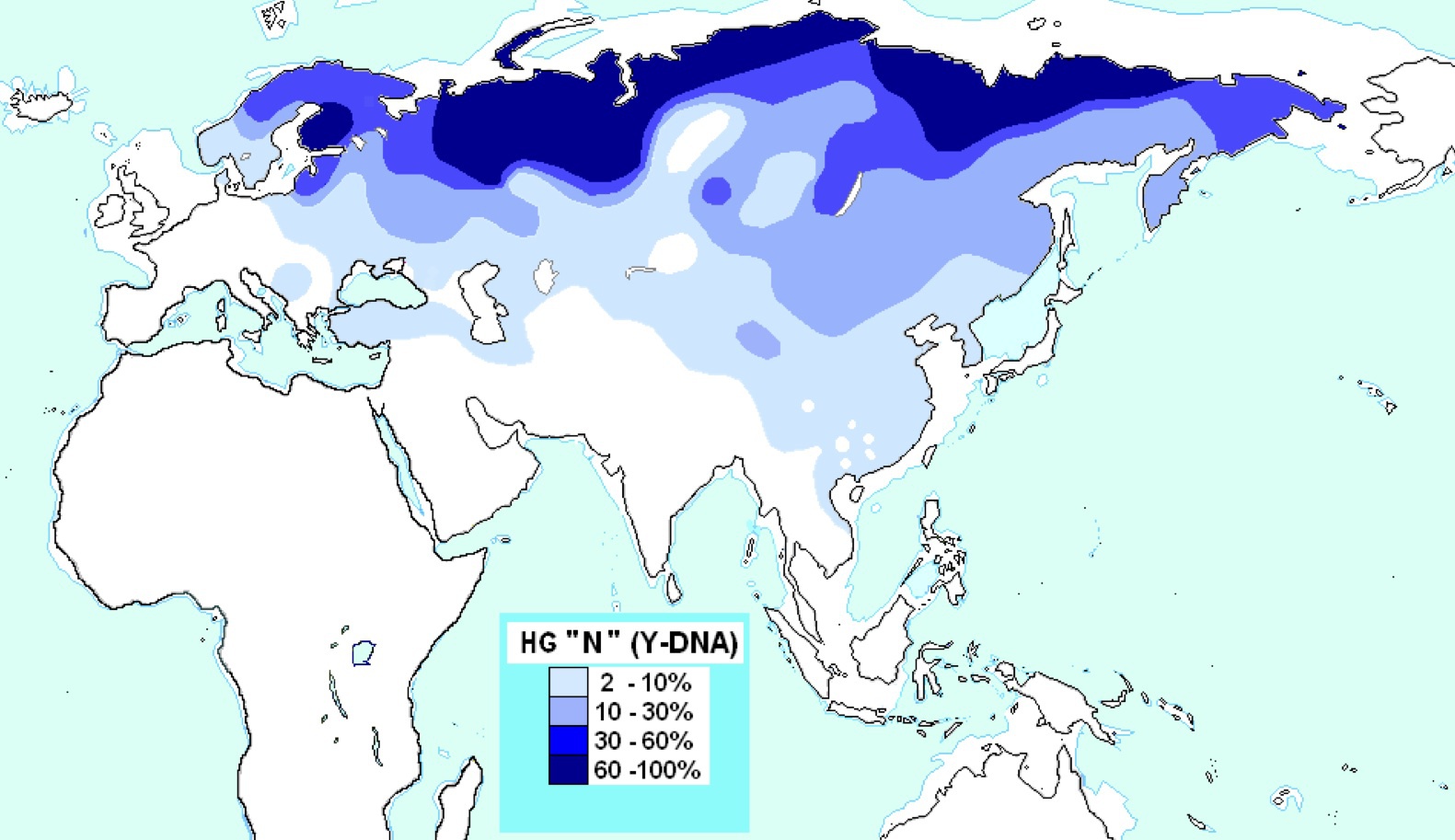
ハプログループNの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループNはフィンランド、バルト三国、ロシア北部からシベリア、北東アジアに広がっている。
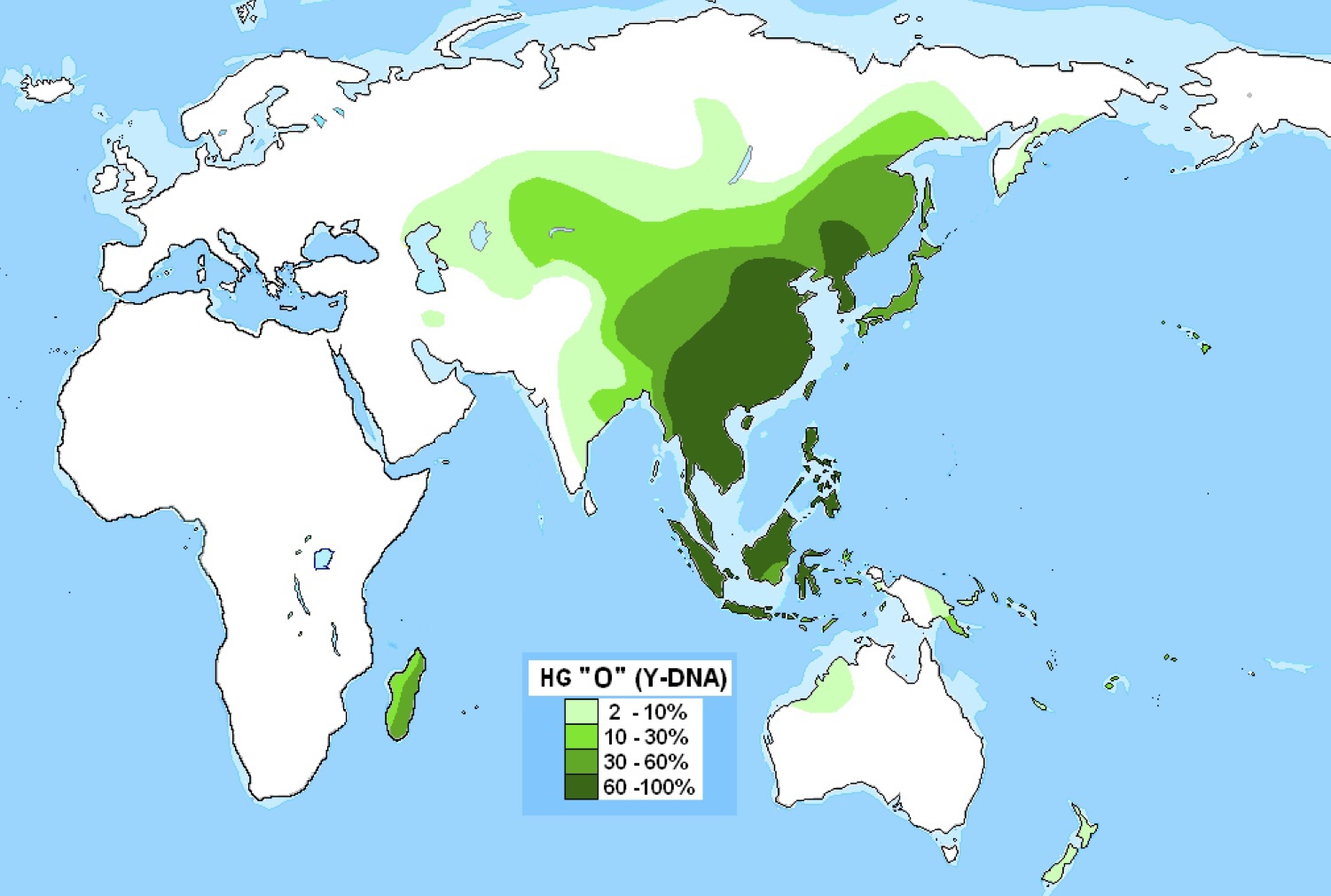
ハプログループOの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループOは東南アジア、東アジアを中心に拡がり、遠くマダガスカルに及んでいる。
ハプログループNとOの直近の共通祖先はNOとされている。おそらくアラル海の東あたりで発生し、その子孫のNが北に行き、別の子孫Oが南に展開したと信じられてきた。だが現在のところ、NOタイプを持った人物はまだ発見されていない。NOの起源については別意見もあり後述する。
(h)ハプログループQとR
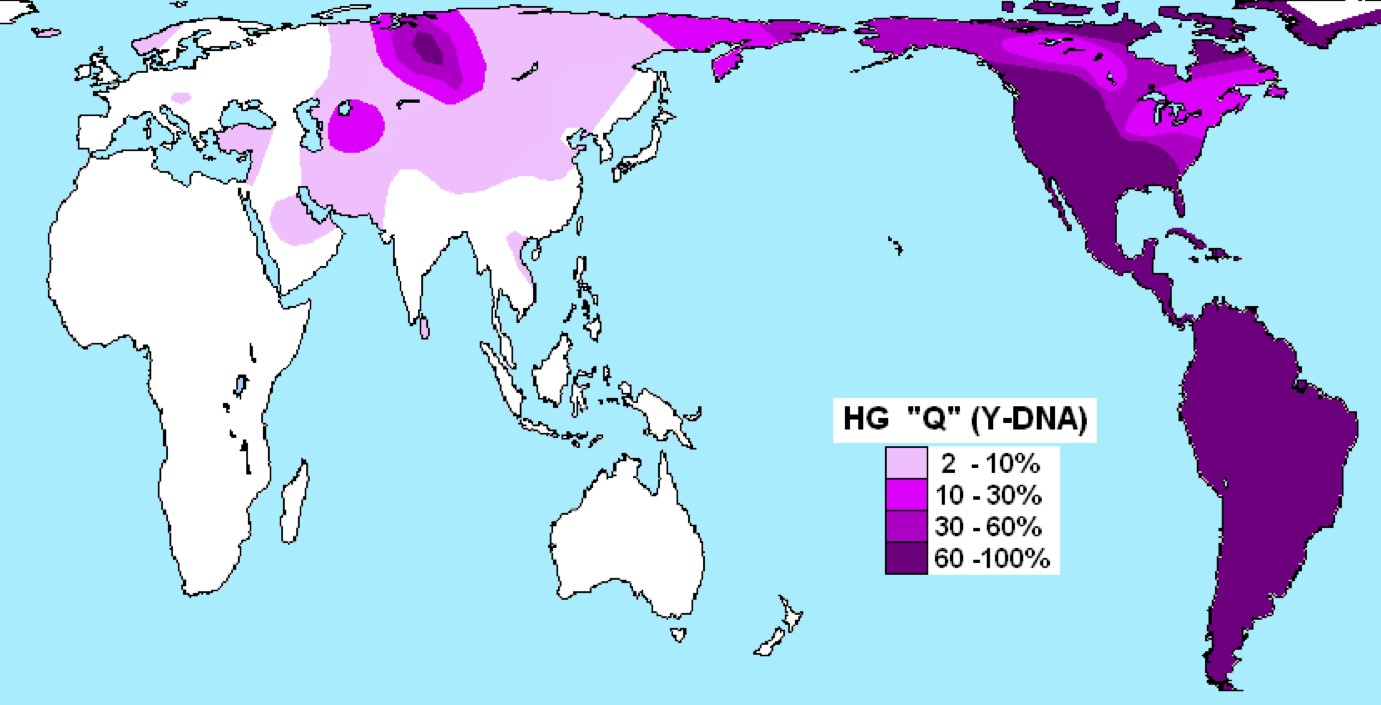
ハプログループQの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループQはシベリア、北アジア、北米、中米、南米と広く分布する。中南米と北米西部南部の先住民の大部分を占める。他の図とあわせ、アメリカ先住民はR、C3、Qのハプログループから構成されていると言って良いことがわかる。
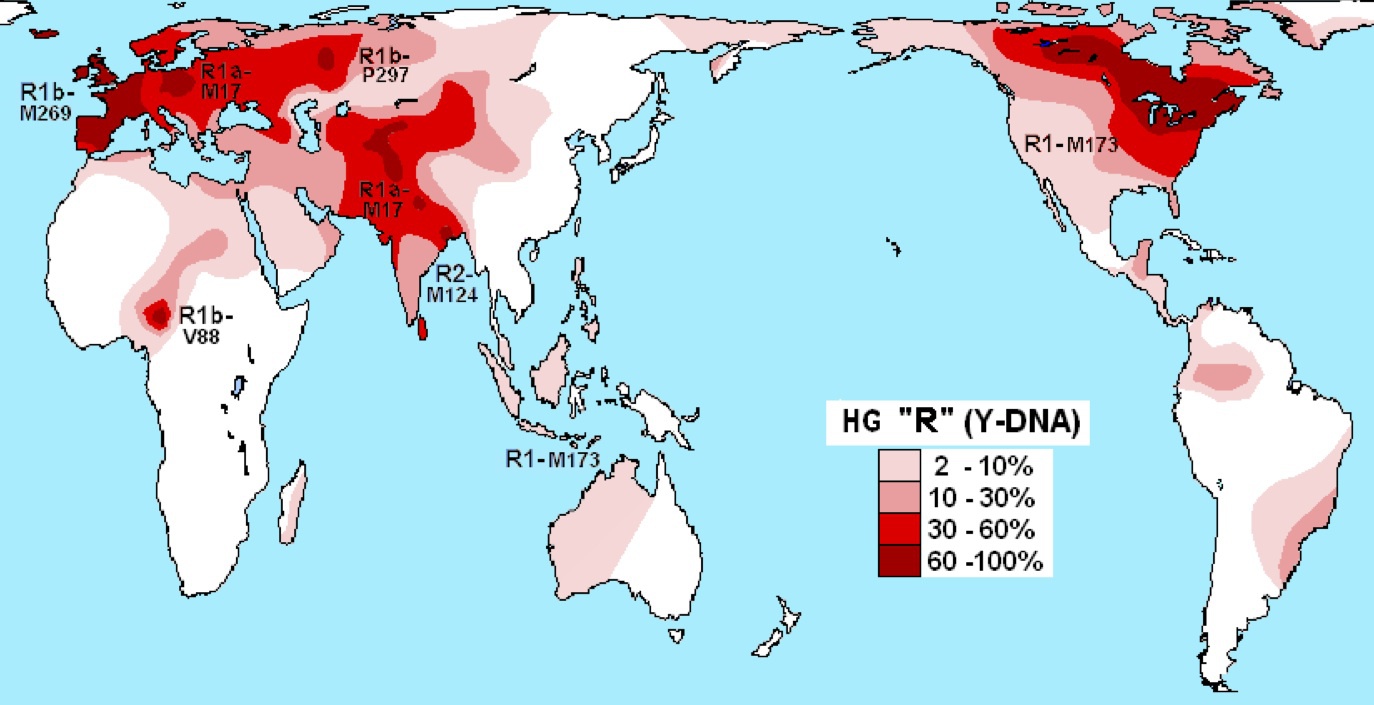
ハプログループRの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループRは西欧、東欧、西ロシア、中央アジア、南アジア、北米など広く分布し、アフリカ、東南アジア、オーストラリア、南米にまで認められる。
(i)ハプログループMとS
ハプログループMの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループMはニューギニア島中央部(西側のイリアンジャヤに多い)を中心としたメラネシアおよび東部インドネシアに分布している。
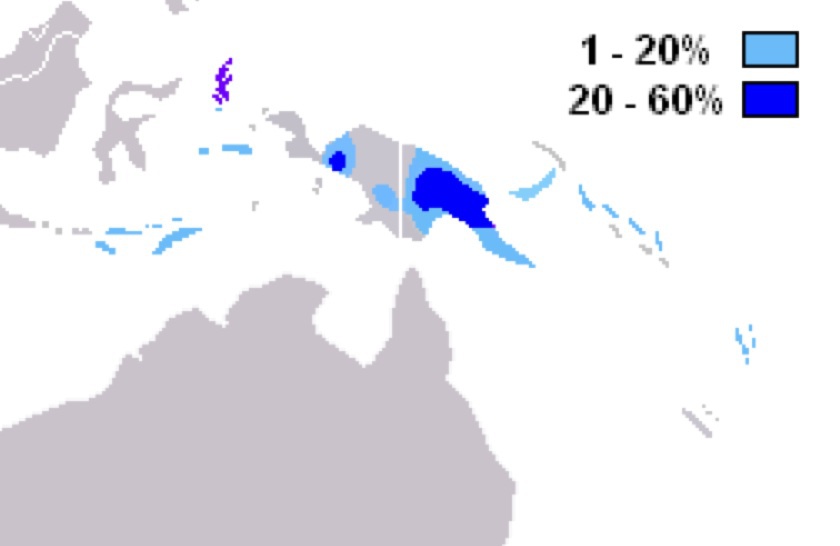
ハプログループSの分布
Wikipediaによる(CC BY-SA 3.0)
ハプログループMと同じくSもメラネシア中心だが、ニューギニア島の東側パプアニューギニアに特徴的である。
ⅲ)各Ydnaハプログループの世界分布概観
以上まとめて全体を概観した図が次である。
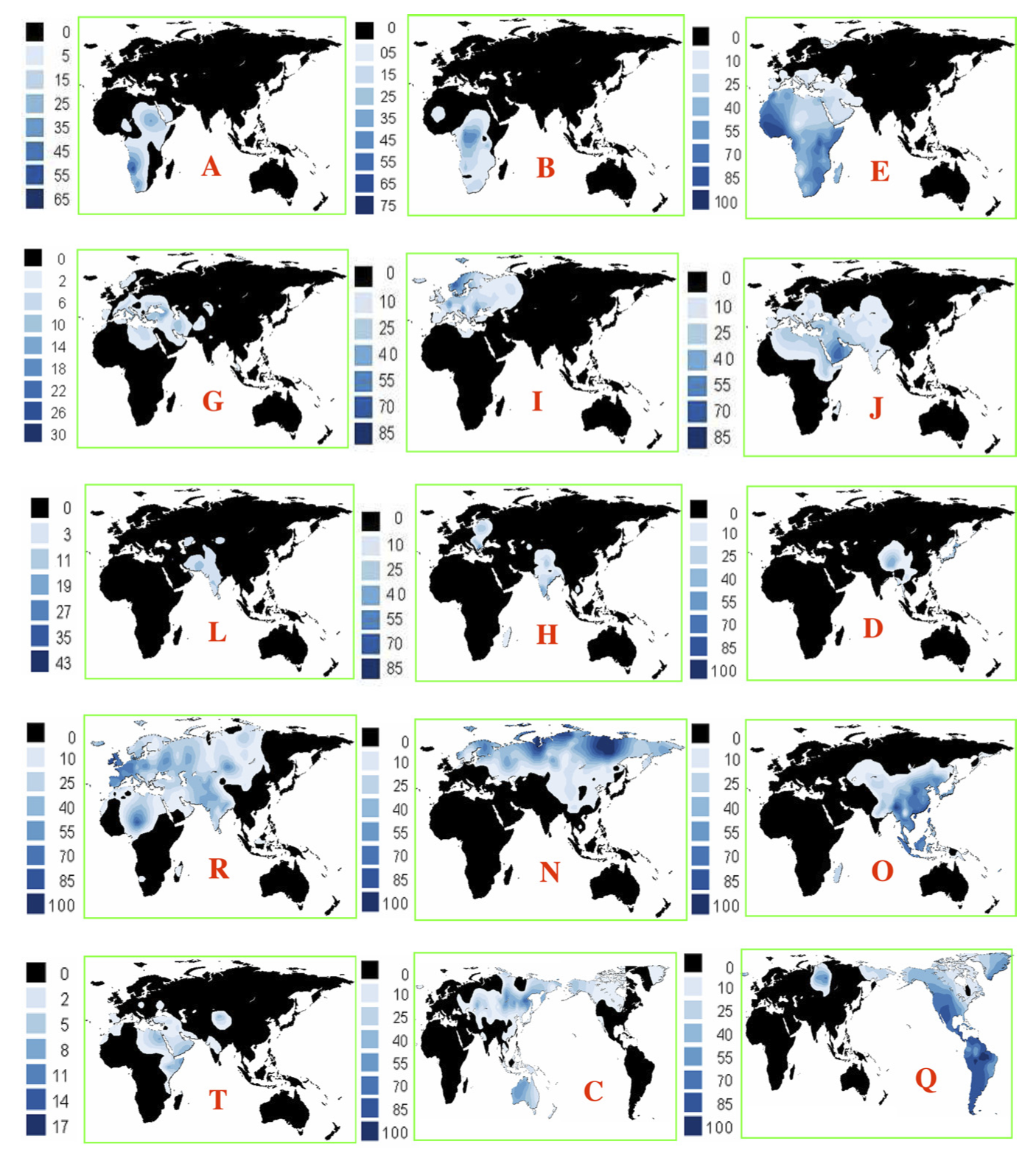
Chiaroni 2009 による。
Ydnaハプログループの系統樹と世界分布地図を組み合わせると次の図のようになる。
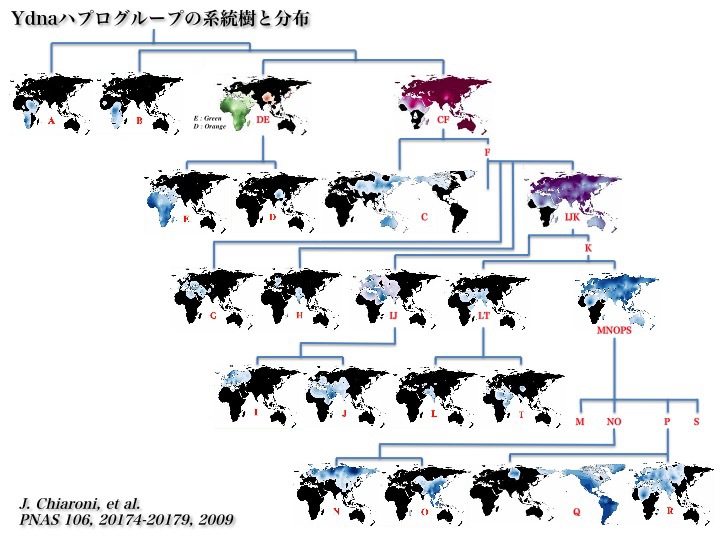
Chiaroni 2009より改変
出アフリカ大陸を果たしたハプログループD、Cが分岐した後、Fの子孫たちが全世界各地に散らばる。Fは現在の人口全体の90%以上を占めるとされる。
そこからG、Hが枝分かれし、IJKからIJを経てIとJが出る。残るKからLTを経てLとTが分岐し、MNOPS(KxLT)となる。MNOPSからMとSが分かれてメラネシアを中心に分布し、残りからNOとPが生じる。
そしてNOからユーラシア大陸の北端全域にまで拡がったNと東アジアを中心に分布したOに分岐し、Pから南北アメリカ大陸を中心とする先住民の祖となったQとヨーロッパを含むユーラシア大陸西側からアフリカ大陸北部地域に至るまでの人々の祖先となったRが出た。
Ⅲ)出アフリカ後の人類推定移動経路
大くくりの分類で移動経路を推定してみよう。
ⅰ)ハプログループCT
ハプログループAとBをのぞく全てのハプログループCTの変異は、アフリカ北東部のどこかで起こり、そのあと一部が出アフリカを果たす。DEに分かれる変異とCFに分かれる変異は、65000年前頃に起こったと推定される。CとDは海岸沿いのルートをたどり、アラビア半島南海岸、西アジア沿岸部、インドの海岸線を経て、東南アジアを含むスンダランドに至ったと考えられる。
ハプログループCは上述の通り、オセアニア、東アジア、北アジアを経て、陸続きだったベーリング海峡を通ってアメリカ大陸に達し各地に展開した。
ハプログループDは北上して日本に至り、そこから陸続きだったユーラシア大陸に渡りチベットにまで達した。当初ひろくシナ大陸に拡がっていたが、O集団により圧迫を受けてシナ大陸中央部からは駆逐されてしまったと考えられている。
ⅱ)ハプログループF(G、H、IJK)
ハプログループFの子孫は全人類人口の90%を占める。全てはサハラ砂漠以南を除く地域に分布している。IJを規定する変異は45000年前頃に起こったと推定され、一団は中東や南アジアを出てヨーロッパに向かった。
約30000年前、中東からパキスタンあたりでハプログループGは生まれ、新石器文化を携えてヨーロッパに向かったと考えられている。Hの変異は30000~40000年前頃にインドで起こり、そこにとどまっただろう。同じ頃に変異して分かれたハプログループKは、広くユーラシア、オーストラリア、南太平洋各地に移動していったと推定される。
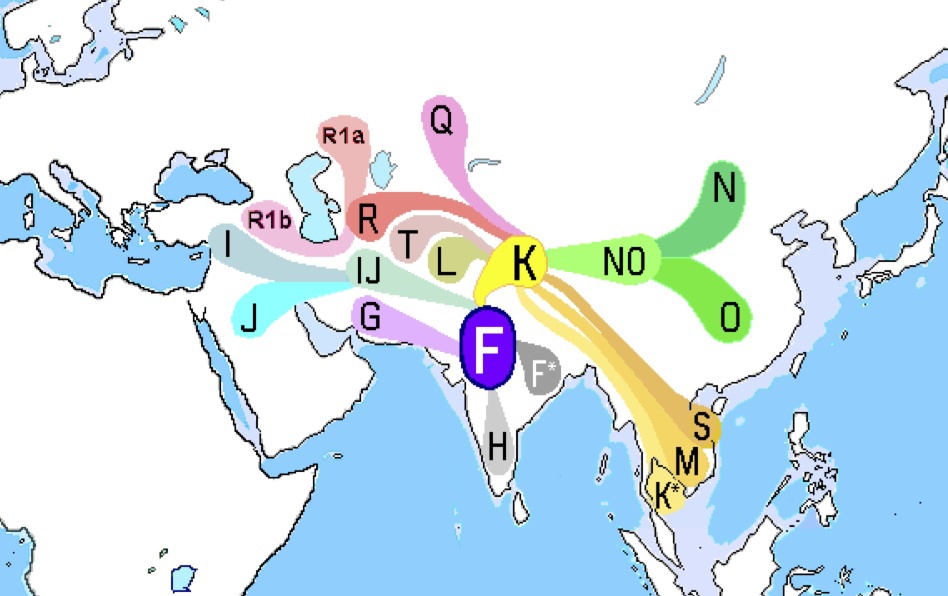
ハプログループFが西アジアから南アジアで生まれ、世界に拡がったという仮説をイラストにしたもの
Wikipediaによる(CC BY-SA 3.0)
ⅲ)ハプログループK(K、LT、MNOPS)
ハプログループKはインド亜大陸、インドネシア、メラネシア、ポリネシア、オーストラリアに見出されたとされている。Lは主に南アジアに認められ、Mはメラネシアに最も多い。
ハプログループNOは約35000~40000年前にアジアで出現したと推定される。Nはユーラシア大陸北端、シベリアからウラル以西に至る人々に最も一般的な集団となった。Oは東アジア、東南アジアに高頻度に見られ、中央アジア、南アジア、南太平洋にも低頻度ながら認められる。
ハプログループPはQとRの祖先だが、Pとしてはほとんど見つかっていない。中央アジアかアルタイ地域に由来するだろう。Qも中央アジアに由来し、発生時期は17000~22000年前。東に向かい北アメリカに至った。
ハプログループRの多くはR1aとR1bからなる。R1aはユーラシアステップに起源を有し、スキタイ文化に関係するとともに原インドヨーロッパ人が増え拡がることにもつながった。基本的に中央アジア、南アジア、東ヨーロッパに見られる。
他方、R1bはおそらく中央アジアを起源としている。西欧で最も優勢なハプログループで、アジア、アフリカの人々にもポツリポツリと見られる。たくさんのサブハプログループに分かれ、その中でも最大の集団R1b1a2は、「現代ヨーロッパ人」特に「西欧人」に最も一般的に認められる。
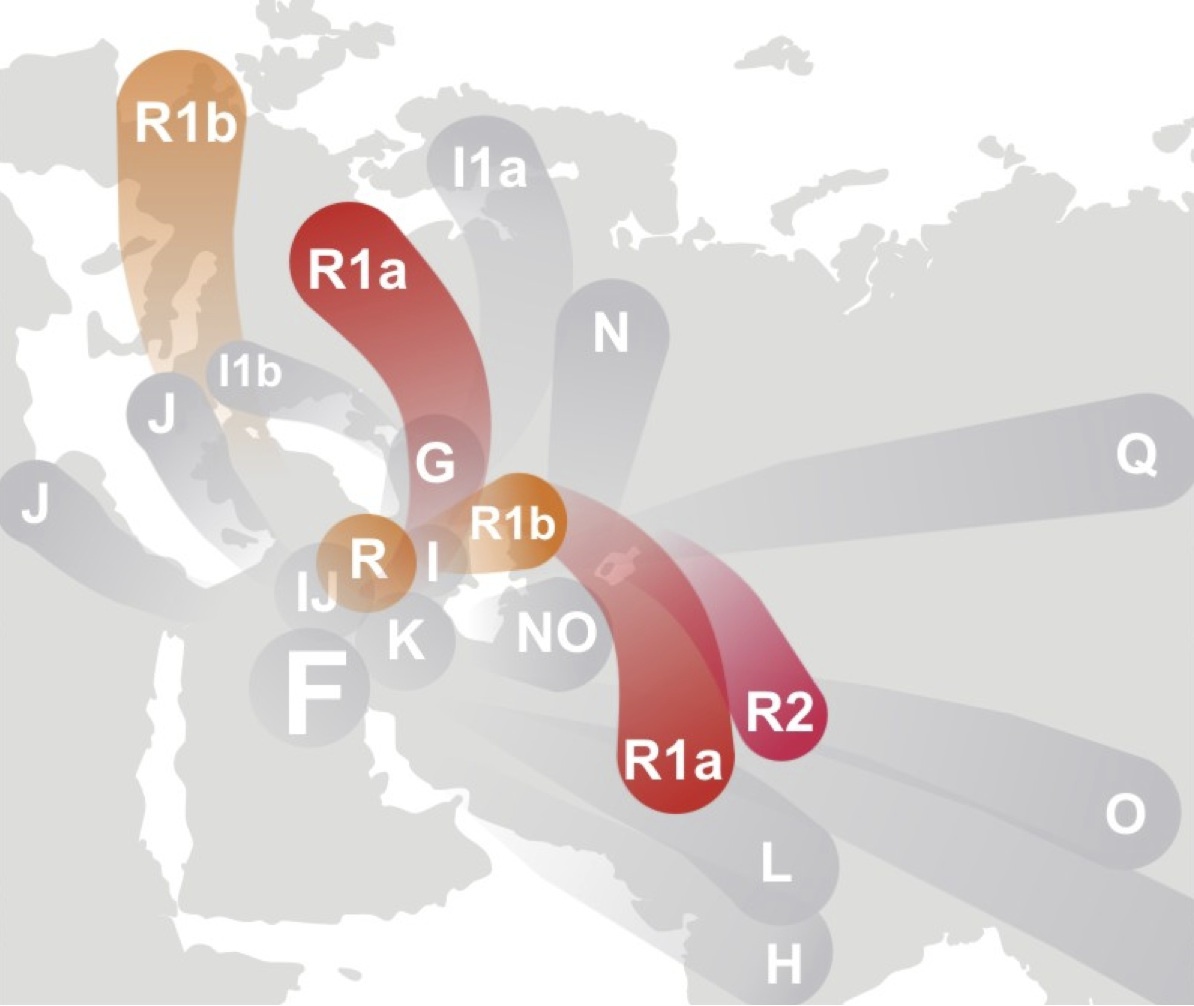
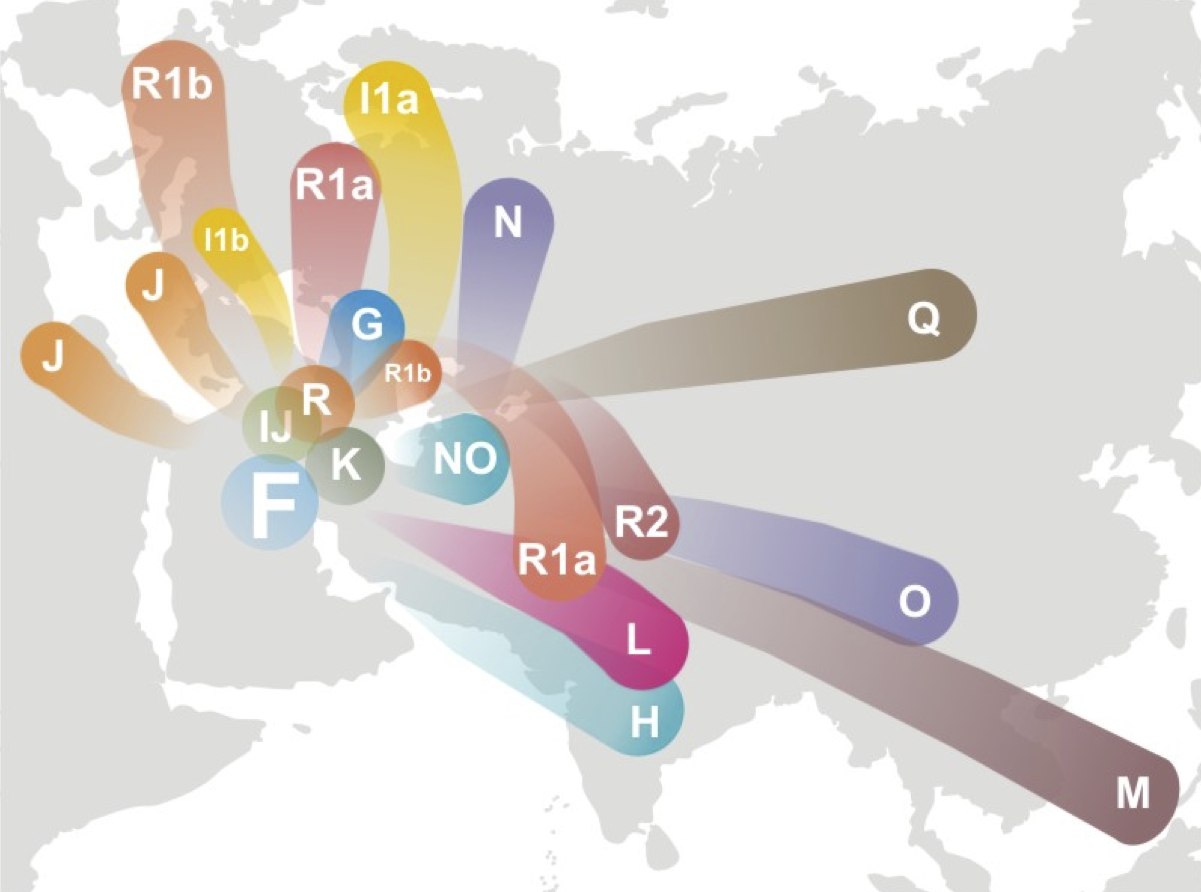
どちらもハプログループFとRが西アジアに生まれ、ヨーロッパとインド亜大陸などに拡がったという仮説をイラストにしたもの
上図はWikipediaによる(CC BY-SA 3.0)
下図はThe Haplogroup F Y-DNA Project by Garland Boyette (http://www.familytreedna.com/public/F-YDNA/default.aspx)による。
Ⅳ)Ydnaハプログループ研究から日本人のルーツを探る前の小さなまとめ
以上のようにYdnaハプログループが報告され、世界における分布が明らかになった。主な分布の様子を別の形でイラストにしたのが次の図である。
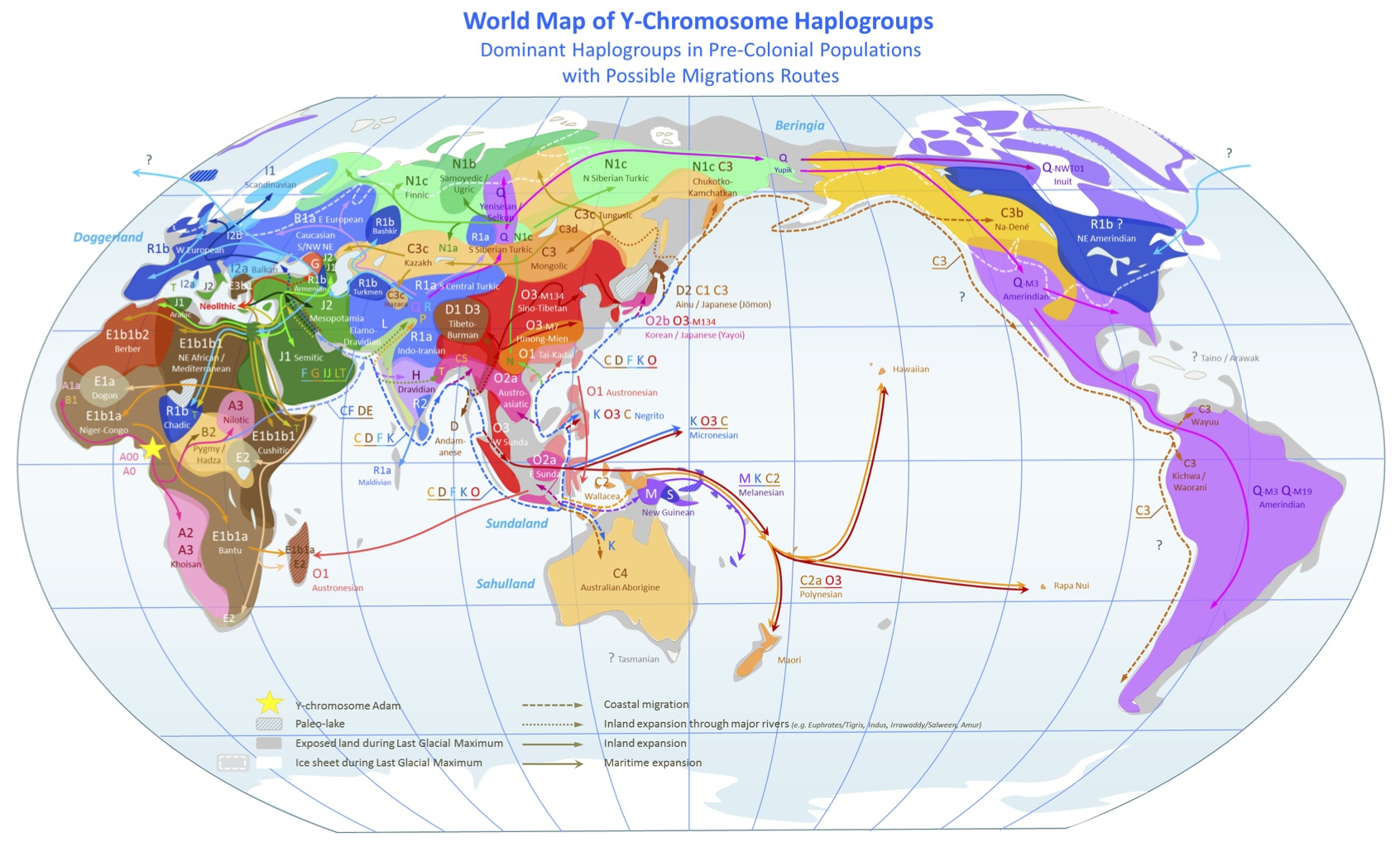
世界各地に存在する主なYdnaハプログループとサブハプログループ
Wikipediaによる(CC BY-SA 3.0)
さらに大きな図はこちら
詳細は触れないが、日本列島はD2/C1/C2が多く分布する北日本、O2b/O3が多い東~西日本、O3が多い九州と色分けされている。ただし図は大まかな特徴付けを表しているのであり、必ずしも正確さを求めてはいない可能性もある。
とはいえ、各ハプログループ移動経路も含めて、本図はたいへん情報量が多く、ずっと眺めていても興味は尽きないほどの傑作イラストの一つと言えると思う。
現代の生きたサンプルでしか解析できないY染色体であっても、ハプログループを追いかけてゆくことで、出アフリカを果たした後の足跡が追えることが判ってきている。これは非常に大きな研究成果だった。インパクトは大きかったのである。
日本人の祖先とは誰か?どのような経路をたどり、いつごろ列島に移り住んだのか?Y染色体DNAハプログループの分布に関する研究から解明しようというのである。
Y染色体ハプログループ全体の詳細系統樹をご覧になりたい方は、ここを参照されたい。
本稿の文献はこちらを参照されたい。
日本人のルーツ(Ⅶ)日本人Y染色体DNAハプログループの特徴
2014.1.19
日本人Y染色体DNAハプログループの特徴 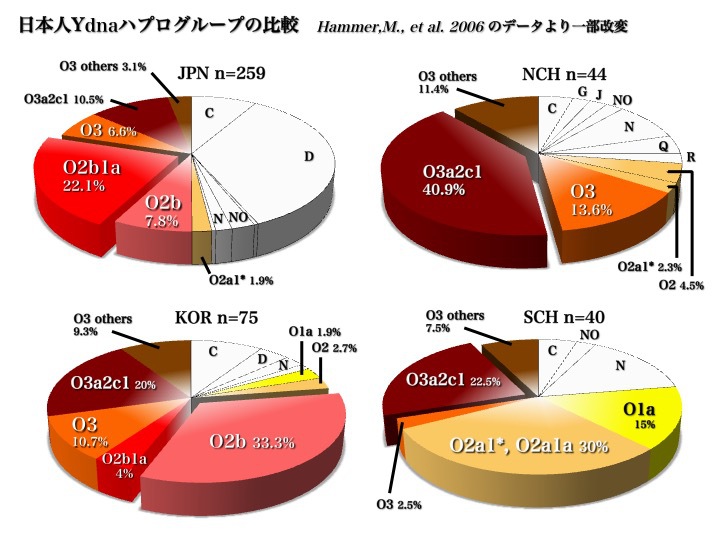
Y染色体DNAハプログループからは日中の共通性は見出せない
本稿の文献はこちらを参照されたい。
日本人Y染色体DNAハプログループの特徴
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
このページのまとめ
1)現代日本人のY染色体DNAハプログループの頻度分布が解析された。
2)メインはC、D、Oの三つである。
3)CはC1、C3、DはD2a1b、D2a*、D*のサブハプログループである。
4)OはO2b、O2b1a、O3a2のサブグループである。
5)日本列島に特徴的なC1は海岸沿いに南方から来た。
6)日本に特徴的なD2も海岸沿いに南方から到達した。
7)O2b*、O2b1aは日本人、朝鮮人に特徴的で、中国大陸には存在しない。
8)シナ大陸の4〜6割のO3a1c、O3a2c1とその類縁グループは、日本に8%しかいない。
9)シナ大陸→日本へ移動したなら、O3a1cとO3a2c1が並存するはず。
10)O3a1cは日本に稀。従ってシナ大陸→日本への移動は限定的。
11)大陸とつながっていた頃、O2bグループは日本列島や朝鮮半島に到達した。
12)大陸との分離後、O2b1aが日本で、O2b*が朝鮮半島で多数派になった。
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
Ⅰ)現代日本人Ydnaハプログループの頻度分布
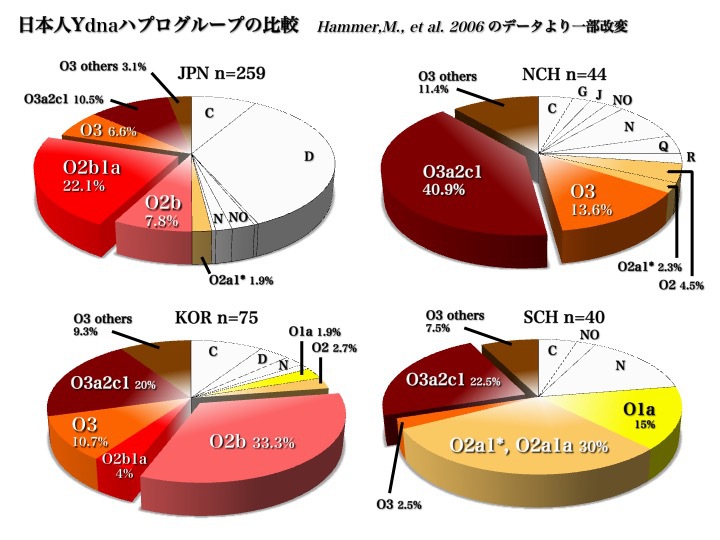
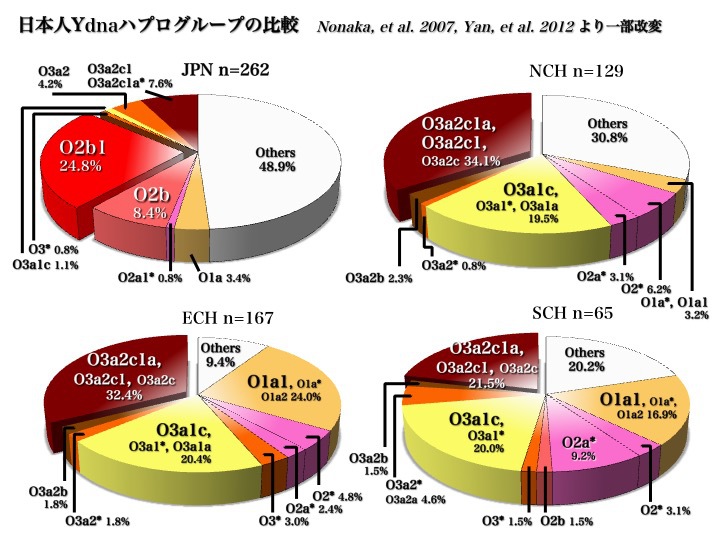
日本人はどこから来たのか?論じる第一歩は、もちろん現代日本人を構成するYdnaハプログループの種類とそれぞれの頻度分布を知ることである。2006年と2007年のデータが大きな傾向を教えてくれる。
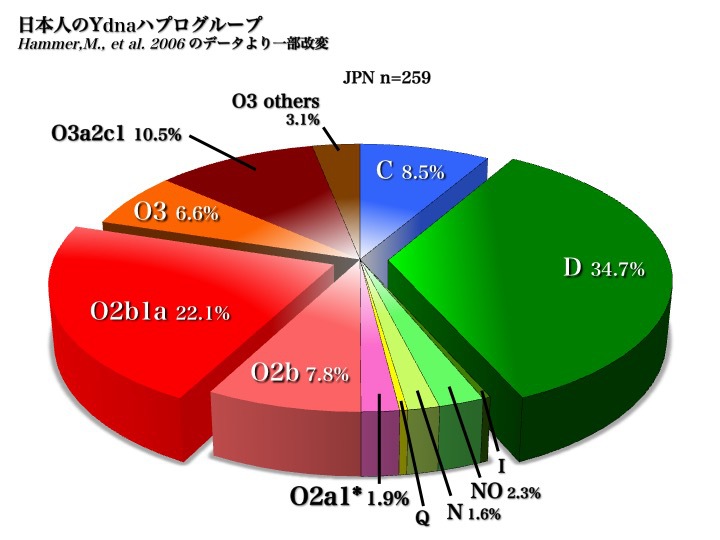
日本人のYdnaサブハプログループ
上図2006年のデータから、日本人はC、D、O(O2b、O3)の三つの集団から成り立っていることがわかる。
また、五つのハプログループD、O2b1、O3a2c1、C、O2b*により、現代日本人の大部分が構成されていると言える(以上Hammer 2006による)。
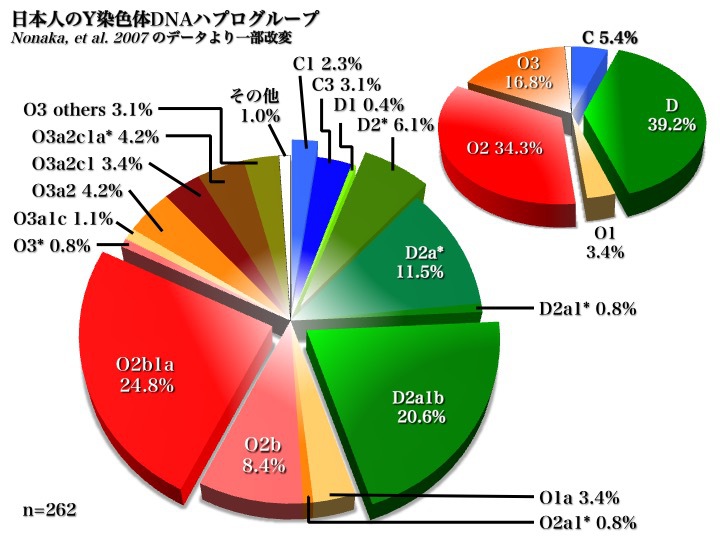
日本人のYdnaサブハプログループ
2007年のデータからも同じ傾向が示されている。上図の右上のパイチャートのようにC、D、O(O2b、O3)の三つから成り立っていることがわかる。
もっと詳しく見ると、D2a1b、D2a*、D2*、O2b1a、O2b*が五つの主要集団であり、O3a2c1、O3a2c1a、O3a2、O1a*、C3、C1が頻度5%未満ながら続く(以上Nonaka 2007による)。
研究の進展によりハプログループC、D、Oはいずれも大きな呼称変更がなされた。もっとも影響を被ったのがO2b1とO2b*である。O1を特徴づける変異MSY2.2がハプログループの基準を満たさないことが判明した。
そのため、O1とO2がともにO1グループに属すことになり、O3はO2に呼称変更された。Oを特徴づける遺伝子変異はM175と呼ばれるマーカーである。ハプログループO-M175と称される。Hammerの研究で用いられた当時のO2関係の遺伝子マーカーはP31、M95、M88、SRY465(M176)、47zである。47zは現在CTS713にとって代わられているが、混乱を避けるためそのまま47zを用いる。
当時の研究で用いられたO2b関連のマーカーはM175/P31/SRY465/47zで、O2b1aはその全部を併せ持つタイプだった。ひるがえってO2b*の方はM175/P31/SRY465を有するけれど47zを欠くタイプだった。こうした遺伝子マーカーを使った方が比較的簡単に表現することが可能で、後者はO2b(xO-47z)あるいはO2b(xO2b1a)と表せる。47zあるいはO2b1aを除くO2bに属するすべてという意味である。
2017年に合わせた表記にすると、O1b2(xO-47z)あるいはO1b2(xCTS713)となるだろうが正確さに欠ける。O-M175>O-P31>O-SRY465>O-47zとO-M175>O-P31>O-SRY465(x47z)あるいは(xCTS713)とした方が当時の研究結果を正確に表現することになろう。
それぞれの集団が系統樹上どの位置に属するかは次に解説する。
Ⅱ)日本にたどり着いたYdnaサブハプログループとその世界分布
ⅰ)日本にたどり着いたハプログループC、D、Oの系統樹
上述のように日本列島にたどり着いたのはC、D、Oであることがわかった。ではそれぞれの集団はどのような仲間がいるのか見てゆこう。
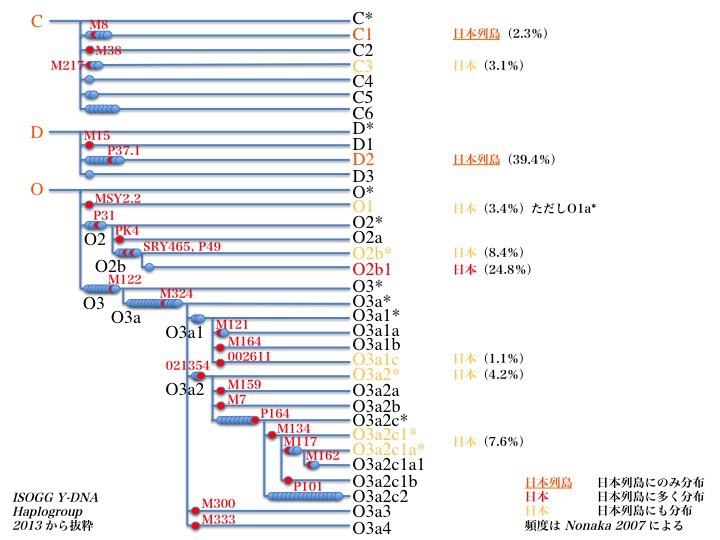
日本にたどり着いたY染色体DNAハプログループC、D、Oと
派生するサブタイプ
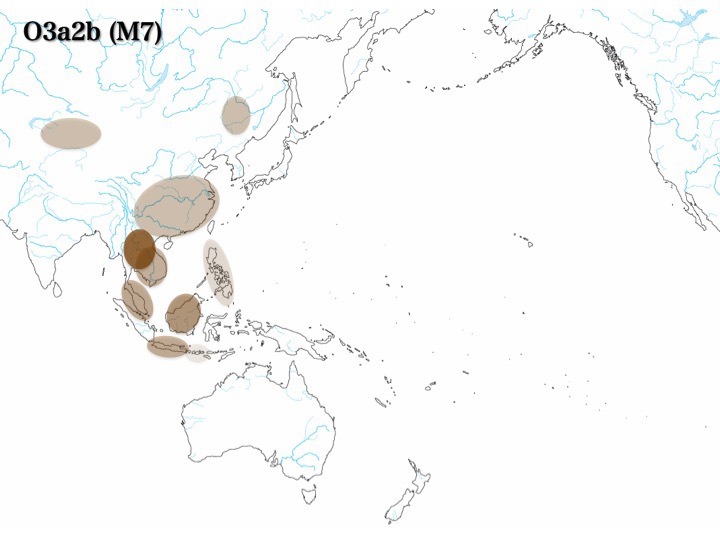
日本にたどり着いた主なY染色体ハプログループC、D、Oそれぞれの系統樹が上図である。ハプログループCはC*/C1/C2/C3/C4/C5/C6の七つのサブハプログループとなる。ハプログループDはD*/D1/D2/D3の四つのサブハプログループに分岐する。
ハプログループOはO*/O1/O2/O3のサブハプログループに分かれ、O2はさらにO2*/O2a/O2b*/O2b1に分岐する。
またO3はO3*/O3a*/O3a1/O3a2/O3a3/O3a4に枝分かれするとともに、分岐したうちのO3a1とO3a2はさらに多くのサブハプログループを従えている。
日本列島にだけ存在するサブハプログループがある。C1とD2である。他の地域では見出せない。主に日本列島に存在する特徴的なサブハプログループO2b1もある。
他、C3、O1a、O2b*、O3a2とO3a1c、O3a2c1の集団も日本列島に一定数みられるサブハプログループである。
ⅱ)ハプログループCの系統樹とその分布、推定移動経路
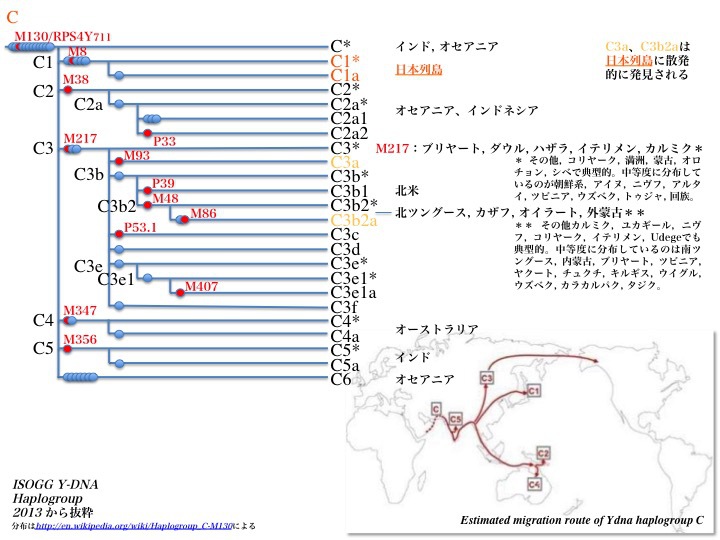
Y染色体DNAハプログループCの系統樹とC集団の世界拡散
ハプログループCは前図で示したC*/C1/C2/C3/C4/C5/C6より、さらに詳しくサブハプログループに分枝している。C3のうち日本列島に分布するのはC3aとC3b2aであり、北米にはC3b1、東アジアやシベリア、シナ大陸にはC3b2、C3b2aおよびその他C3サブハプログループ集団がいる。
オーストラリアに移動したのがC4、インドに留まったのがC5、オセアニアまで行ったのがC6と多岐にわたっている。他にインドにとどまったりオセアニアまで行ったりしたのがサブハプログループC*である。
北東アジアに分布した集団は規模は大きく、満洲族の26.9%からオロチョン族の90.9%にまで至る。モンゴル族でも52.3%に及ぶ(Hammer 2006による)。
推定されているハプログループCの世界移動経路が図右下に示されている。正確を期すならば、インドを出た後もう少し海岸線沿いにルートをとって東南アジアを通りながら北上し、日本列島にたどり着いたと考えられる。
日本人がモンゴル系などと言われたり、北方から日本列島に渡って来たとされたりすることについてコメントするなら、次の何点かを指摘せざるを得ない。
1)日本人におけるCの割合いは5.4~8.5%とそれほど高くない。
2)その中でも、日本人にしかいないC1が2.3~5.4%を占める。
3)C1は九州をのぞく全国(沖縄から青森)に分布し、特に北に多い傾向は示さない。
4)北方系部族に多いC3は、アイヌに見出される他、逆に西日本に多く分布する。
これらは、たといハプログループCに限ったとしても、日本人が単純にアルタイ系、ツングース系またはモンゴル系であるなどとは決して言えないことを示唆している。
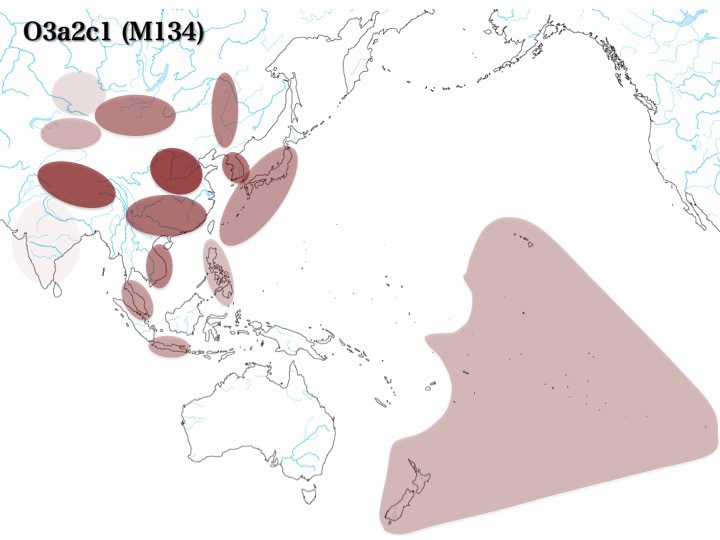
ⅲ)ハプログループDの系統樹とその分布、推定移動経路
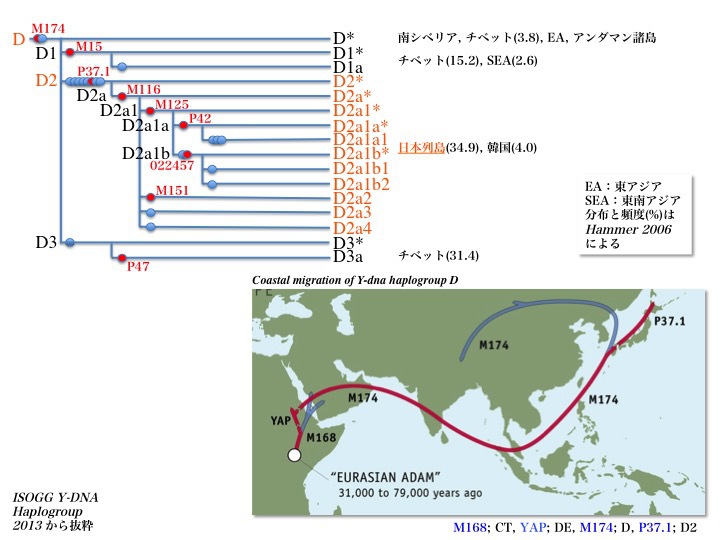
Y染色体DNAハプログループDの系統樹とDの推定移動経路
ハプログループDはD*、D1、D2、D3の四つに分かれる。D*は南シベリア、チベット、北東アジア、インド洋東側に浮かぶアンダマン諸島に見られる。
そのうちD1はチベットに多く(15.2%、Hammer 2006)、東南アジアにも少数(2.6%)分布している。D2は日本列島にのみ存在が確認され(15.2%、同左)、韓国にも4%ほど認める。他方、D3はチベットで大きな集団を形成している(31.4%、同左)。
ハプログループDは、図右下のような移動経路が推定されている。東インド洋のアンダマン諸島に留まったグループ、日本を経由して南シベリア、北東アジア、チベットまで移動したグループは親ハプログループD*のままである。
経由中の日本で枝分かれし、日本列島にとどまって沢山ふえ拡がったのがD2サブハプログループである。恐らく枝分かれした子孫が日本にとどまり、枝分かれしなかった残りが旅を続けてチベットにたどりついたと思われる。
日本を経由して大陸に渡った親ハプログループから、別途枝分かれしてチベットにたどり着いた一団もいる。それがD1、D3サブハプログループである。
日本列島を経由しない経路を想定する人もいる。東南アジアからビルマのエーヤワディー川やタイのチャオプラヤー川を遡ってチベットに行き着いたのかもしれない。
黄河か長江を遡ってチベットにたどり着いたか、それともビルマやタイ経由だったのか?どちらにしても、海岸線をつたって移動するのとどちらが困難な旅だっただろうか。
ハプログループDが一時期シナ大陸に広く存在していた可能性を指摘する見解もある。その後優勢になった別のグループからの圧迫を受け、周辺地域であるチベットや東アジア、南シベリアに追いやられたとの立場をとる。
ⅳ)ハプログループO1、O2、O3の系統樹とその分布
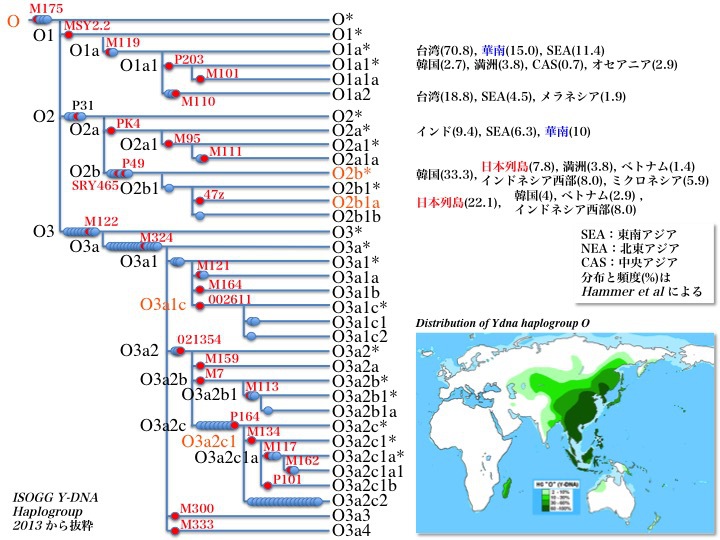
Y染色体DNAハプログループO1、O2の系統樹とそれぞれの分布地域
ハプログループOはインド、東南アジア、東北アジア、華南、華北、朝鮮半島、日本列島に広く分布している。アジアを特徴づけるハプログループである。
サブハプログループO1/O2/O3はさらに幾つにも分岐していった。O1は五つ、O2は八つ、O3は二十一のサブハプログループに分かれている。
それぞの分布を順を追って解説してゆく。
1)O1は特に台湾に多く、華南、東南アジアに拡がっている。
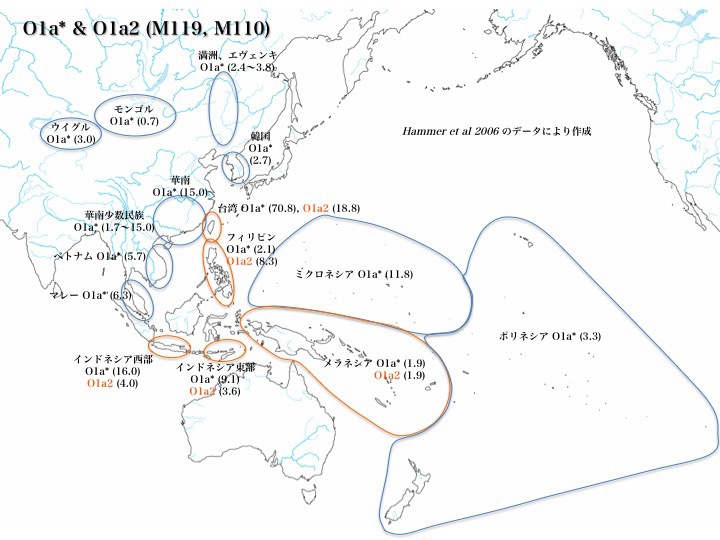
ハプログループO1の分布
台湾原住民は(台湾では「先住民」というと既に滅びて今は存在しない部族を意味する)、O1a*とO1a2*で約九割に至る。O1は実に台湾原住民のハプログループなのである。
他にO1a*はインドネシアで9.1~16.0%、ミクロネシア、華南でそれぞれ11.8%と15.0%、一部華南少数民族でも15%に達する集団がいる。O1a*は広く東南アジア、東アジア、北東アジア、蒙古、ウイグル、オセアニアに分布する。
また、O1a2*の分布はO1a*分布域の中央部に限定される。台湾、フィリピンにそれぞれ18.8%と8.3%を占めるのが特徴的だが、他は4%以下である。O1a2*の方がO1a*より多いのはフィリピンだけである。
日本はどうか。Nonakaらのデータ(2007年)ではO1a*を3.4%認めるものの、Hammerのデータ(2006年)をもとにした本図では分布を認めない。O1は大きな集団とは言えない。
2)O2aはインド、東南アジア、華南に分布する。他方O2bは、主としてO2b*が朝鮮半島に、O2b1aが日本列島に分布している。O2b1aは朝鮮半島とインドネシア西部とベトナムにも存在している。O2b*は日本、インドネシア西部、ベトナム、ミクロネシアにも認められる。
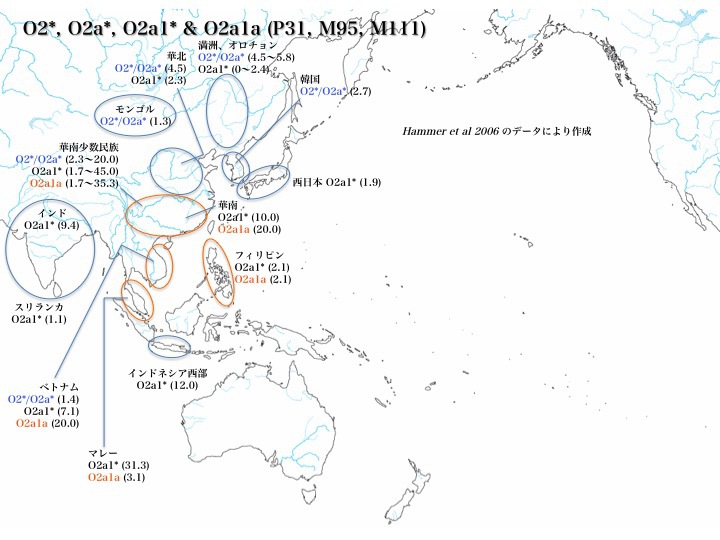
ハプログループO2*、O2a*、O2a1*、O2a1aの分布
南から北へ、O2*、O2a*はベトナム、華南少数民族、華北、韓国、モンゴル、満洲、オロチョン族に分布する。O2a1aはマレー、ベトナム、フィリピン、華南少数民族に分布する。O2a1*はモンゴル、韓国を除くO2*、O2a*分布域と、西日本、インドネシア西部に分布する。
華南少数民族(20.0~35.3%)、華南(10.0、20.0%)、ベトナム(~20.0%)、マレー(~31.3%)に多く、インドネシア西部にも12%存在する(いずれもHammer 2006による)。
日本での分布で見るなら、Hammerのデータ(2006年)でO2a1*が1.9%存在するが、Nonakaのデータ(2007年)ではO2*が0.8%となっている。日本におけるO2*、O2a1の存在比率はごくわずかである。
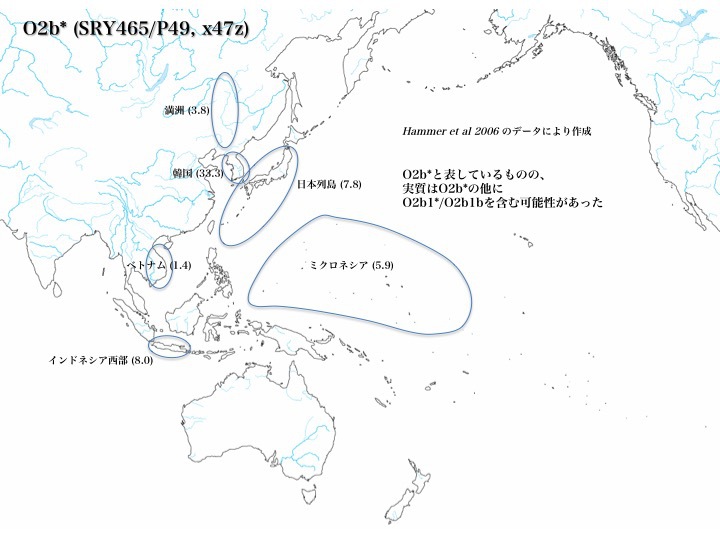
ハプログループO2b*の分布
ハプログループO2b*は韓国に最も多く(33.3%)、次いで日本(7.8%)とインドネシア西部(8.0%)、さらにミクロネシア(5.9%)、満洲(3.8%)、ベトナム(1.4%)と分布している(いずれもHammer 2006より)。
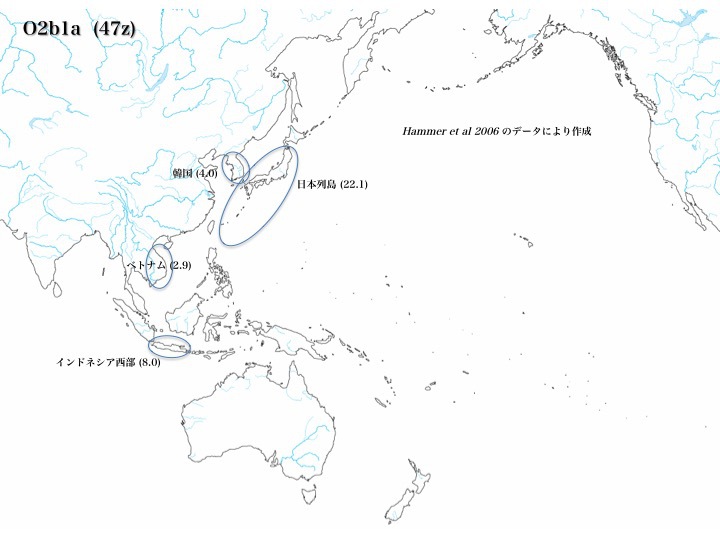
ハプログループO2b1aの分布
ハプログループO2b1aは日本に最も多く(22.1%)、O2b*と同様にインドネシア西部(8.0%)とベトナム(2.9%)、韓国(4.0%)に認められる(いずれもHammer 2006より)。
3)ハプログループO3の系統樹とその分布
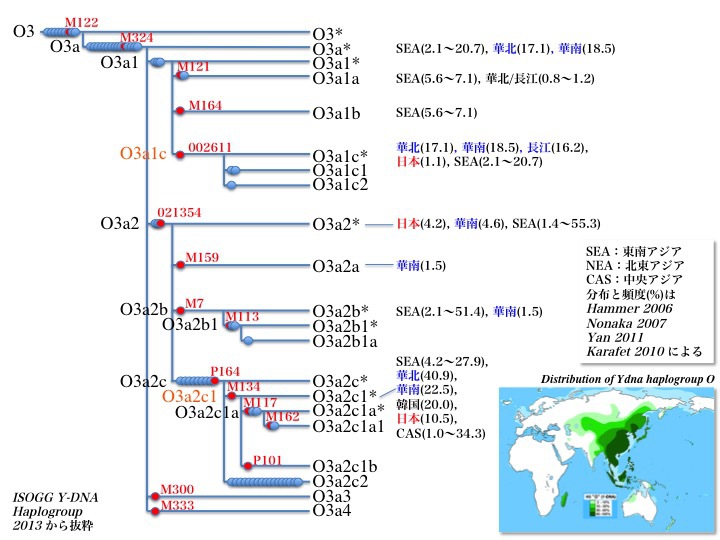
YdnaハプログループO3の系統樹とそれぞれの分布地域
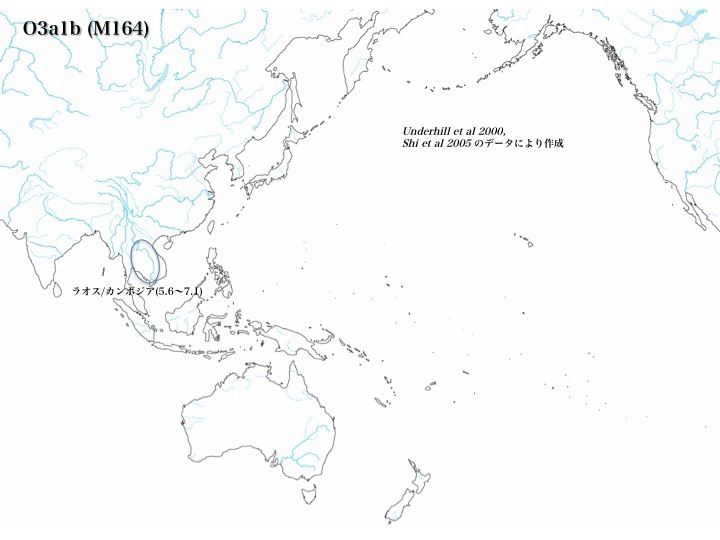
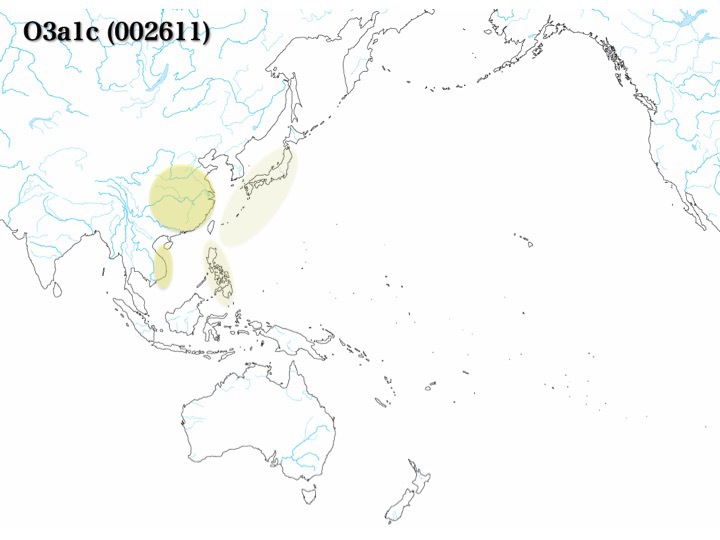
次からの図でも示すようにO3a1a、O3a1bはラオス/カンボジアに存在する。しかしそれ以外は稀で、O3a1aが華南、長江下流域にわずか認めるだけである。対してO3a1cは東南アジアと華南、華北に多く(16.2~18.5%)、日本では3.4%が見つかっている。
ハプログループO3のサブクラスO3aは主にO3a1とO3a2に分かれる。それぞれ順に解説する。
a)O3a1集団はO3a1a、O3a1b、O3a1cに枝分かれする。
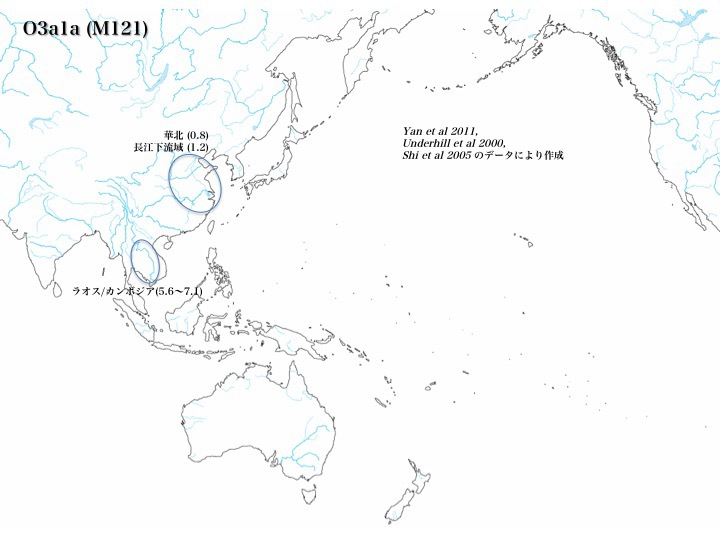
ハプログループO3a1aの分布
ハプログループO3a1bの分布
ハプログループO3a1aとO3a1bは非常に稀である。ラオス/カンボジアに5.6~7.1%程度存在するものの(Underhill 2000、Shi 2005による)、他はO3a1aを華南、長江下流域にわずか認める(0.8~1.2%、Yan 2012による)だけである。
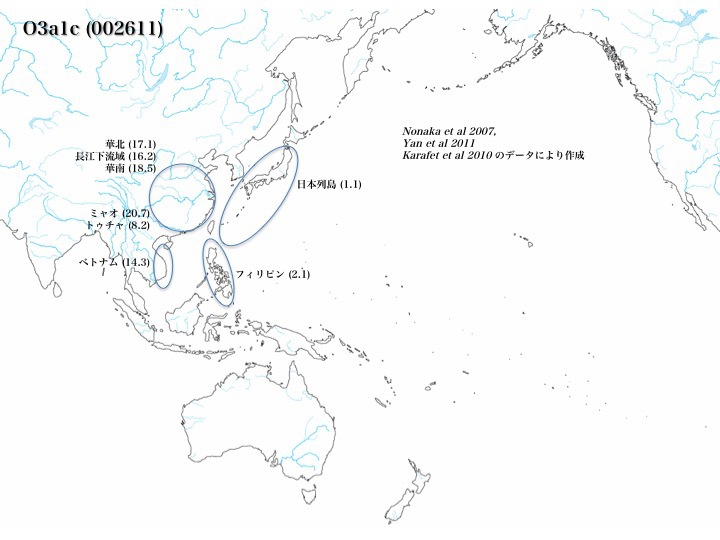
ハプログループO3a1cの分布
それに対してO3a1cはミャオ(20.7%)、トゥチャ(8.2%)、ベトナム(14.3%、以上Karafet 2010より)、華南(17.1%)、長江下流域(16.2%)、華北(18.5%、以上Yan 2011より)に多い。
日本では少数(1.1%、Nonaka 2007)見つかっているだけであるが、東南アジア、シナ大陸では15~20%と充分なプレゼンスを示している。
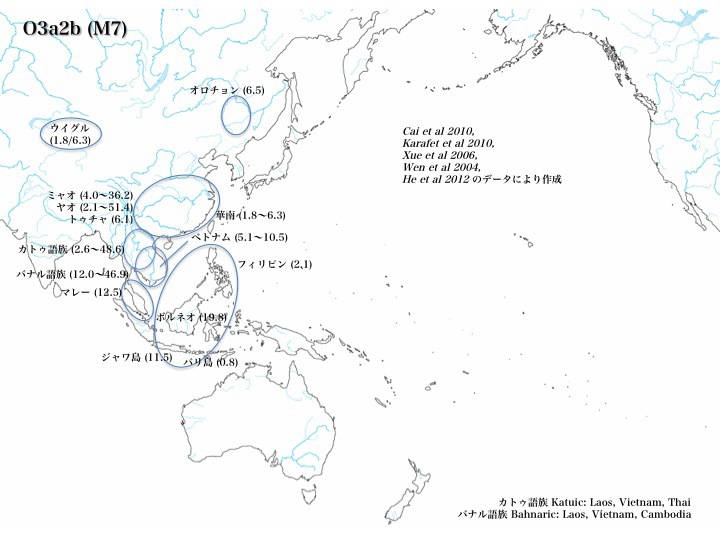
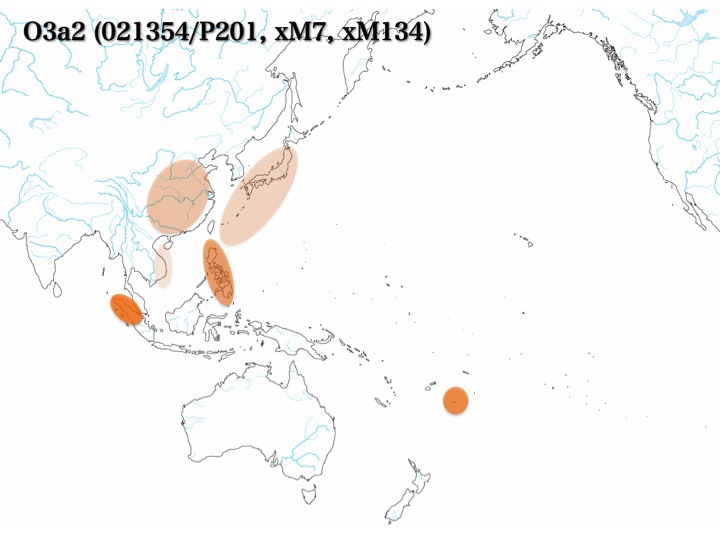
b)O3a2集団はO3a2*、O3a2a、O3a2b、O3a2cに枝分かれする。
ハプログループO3a2集団のうち、O3a2*はフィリピンに多く、ほか東南アジア、華南、華北、日本に1~9%程度見られる。しかしO3a2aはどの地域でも見つかっていないか稀である。O3a2bはボルネオ、ジャワ、マレー、ベトナム、華南やインドシナの山岳少数民族に多く、フィリピンや華南にも認めるものの、日本列島では見つかっていない。O3a2c1は華北、華南、チベットに多く、次いでベトナム、マレー、モンゴル、満洲、韓国に見られる。日本列島にも10%程度分布する。
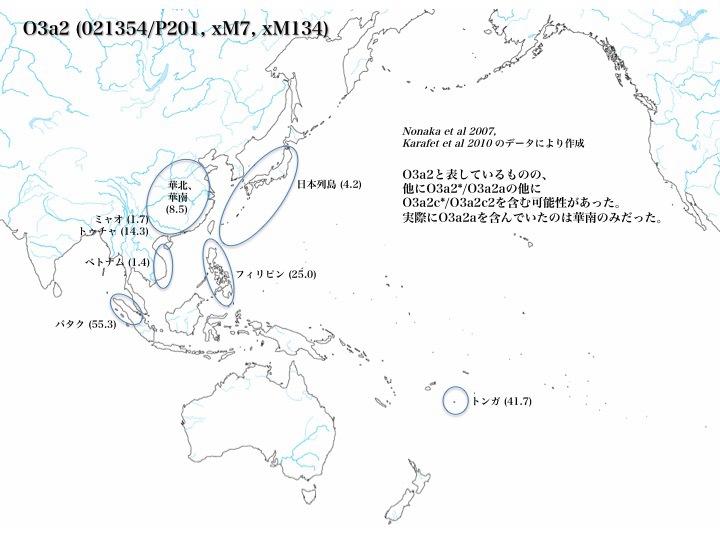
ハプログループO3a2の分布
ハプログループO3a2*はフィリピンに多く(25.0%)、ほかベトナム(1.4%)、華南と華北(8.5%)、少数民族のトゥチャ(14.3%)とミャオ(1.7%、以上Karafet 2010より)、日本(4.2%、Nonaka 2007より)に見られる。
あとKarafetは2010年にスマトラ島北部の少数民族バタクとポリネシアのトンガでそれぞれ55.3%、41.7%の高率でO3a2*を認めると報告した。
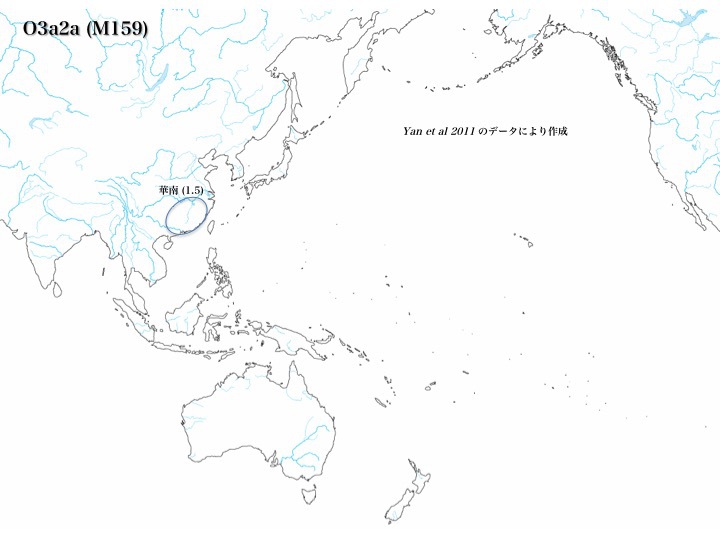
ハプログループO3a2aの分布
ハプログループO3a2aは今のところどの地域にも見当たらず、唯一華南で1.5%程度が報告されている(Yan 2012)。
ハプログループO3a2bの分布
ハプログループO3a2bはボルネオ(19.8%)、ジャワ(11.5%)、マレー(12.5%)、ベトナム(~10.5%)、華南やインドシナの山岳少数民族(~36.2%、~51.4%)に多く、フィリピン(2.1%)や華南(1.8~6.8%)にも認める。しかし日本列島では全く見つかっていない。データは、Cai et al 2010、Karafet et al 2010、Xue et al 2006、Wen et al 2004、He et al 2012による。
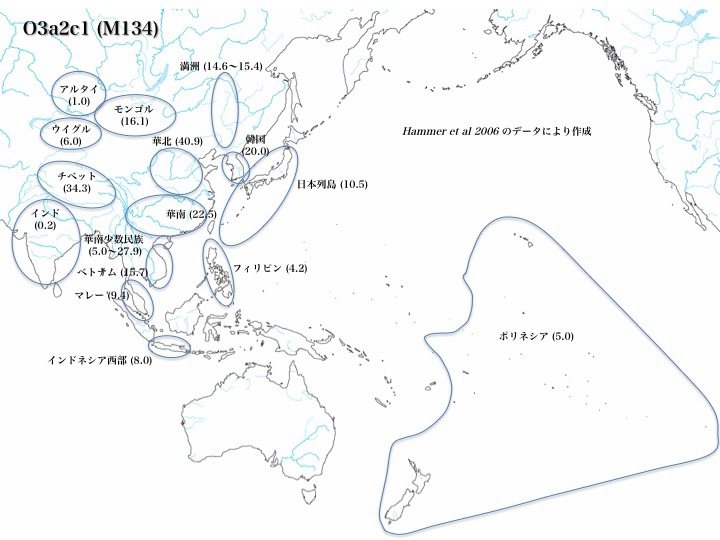
ハプログループO3a2c1の分布
何と言ってもO3の中の最大集団はO3a2c1である。O3a2c1は華北(40.9%)、華南(22.5%)、華南少数民族(~27.9%)、チベット(34.3%)に多く、次いでベトナム(15.7%)、マレー(9.4%)、インドネシア(8.0%)、モンゴル(16.1%)、満洲(15%前後)、韓国(20.0%)に見られ、日本列島にも10%程度分布する。フィリピン(4.2%)の他、ポリネシア(5.0%)、ウイグル(6.0%)、アルタイ(1.0%)、インド(0.2%)など遠く周辺にも拡がっている(いずれもHammer 2006による)。
ⅵ)ハプログループOの分布概観
日本人を構成するハプログループC、D、Oのうち、CとDは移動経路がほぼ推定されている。しかしO集団については分布そのものが非常に複雑で、全体像がとらえにくい。
まず構成する集団の規模から言うと、O集団は次のように類別してよいだろう。
1)O1aまたはO1a2
2)O2a1*
3)O2b集団(O2b*またはO2b1a)
4)O3a2*
5)O3a2c1とその下位集団
赤で示したO1aまたはO1a2、O2a1、O2b、O3a2c1にO3a2c1aを加えて、ハプログループOの系統樹と分布を概観するのに便利なのが次のイラストである。
Y染色体DNAハプログループOの系統樹と分布
一見して、概ね東南アジアや華南を中心に拡がっている傾向が印象的である。O1a、O1a2、O2a1はマダガスカルにまで拡がっており、東南アジアがあくまで分布の中心。例外はO2b集団とO3a2c1aである。
まずO3から見ると、O3は東南アジアにも分布しているが、重心は東アジアから中央アジアにかけての、広くそしてより北側の地域にあるように見える。
一方、O2b集団は中心が日本と朝鮮半島にある。O2bは朝鮮半島で発生したと主張する人々がいる。しかしインドネシア西部とベトナムにも見つかる事実をどうとらえるのか。日本と朝鮮半島附近で発生し、なぜ東南アジアの特定の地域だけに移動したのか。まったく説明がつかない。
東南アジアで発生したO2b*とO2b1aが長い旅をして日本列島と朝鮮半島近辺にたどり着き、一方は日本列島を中心に他方は朝鮮半島を中心に増え拡がったと考える方がよっぽど自然である。
ⅶ)ハプログループOの頻度分布の詳細比較
ハプログループOが日本と朝鮮半島、華北、華南でどのような傾向をもって分布しているか、二つの比較を紹介する。一つはHammer 2006のデータによる比較で、もう一つはNonaka 2007とYan 2012のデータを比べたものである。
日本人Ydnaハプログループの頻度比較
日本人Ydnaハプログループの頻度比較
どちらのデータからも、日本人が朝鮮半島の人々ともシナ大陸の人々とも明らかに違うことがわかる。まずO集団の割合である。日本では五割をわずかに超える程度であるのに対し、華南、華北、韓国いずれも七~八割に達する(Hammer 2006)。
シナ大陸ではO3a1c、O3a2c1とその類縁集団が全体の四割から五割を占めるのに対し、日本ではそれらは10%に満たない。またO1グループとO2aグループがシナ大陸では12.5~31.2%程度存在するのに対し、日本では3%ほどしかいない(Nonaka 2007、Yan 2011)。
日本と韓国はO2b1a、O2b*が多く著しい特徴を共有している。もっとも日本ではO2b1a>O2b*であるのに対し、韓国ではその逆でO2b*>O2b1aという比率である。他方、華南にも華北にもO2b集団は認めない(Hammer 2006)。
華南と華北を比べると、二つの比較で大きな違いがある。Hammer 2006のデータでは、O2a1、O2a1aが華南最大の集団でそれにO1aが加わって半数近くを占めるのに対し、華北最大の集団はO3a2c1でそれにO3*とした集団が加わって半数以上になる。
他方、Yan 2011のデータでは、華南でO2a1、O2a1aの集団が三割を占めるなどのHammer 2006に見られたような特徴はない。これは調査対象とした母集団の違いを反映しているのかもしれない。一つ言えることは、シナ大陸は地域差が大きく均一な集団では決してない、ということである。
ちなみにO以外のハプログループを見ると、日本ではDが圧倒的でCがそれに加わりほとんどを占めるのに対し、韓国ではCが多く、残りはD、Nと続く。華南、華北ではNが最も多く、続いてC。華北の特徴としてG、J、NO、Q、Rが混じっていることがあげられる(Hammer 2006)。
これらからどんなことが言えるか?日本を含む東アジアの地域はOという最大のハプログループを共有しているものの、人々が一方から他方に流れたと推定できる単純な傾向はないということだ。
ただ、日本と朝鮮半島のつながりが、O2b1aとO2b*という両国に特徴的な集団の存在からうかがえる。しかし両者の交流を、いわゆる弥生時代以降と歴史時代に限定して良いのだろうか?もっと前の交流を無視して大丈夫なのだろうか?
たとえば、日韓合邦時代の35年間に最大で約200万人が朝鮮半島から日本に流入した。それは朝鮮半島の人口の7.7%であったが、日本では2.7%程度と見積もることができる。ここ二千数百年ほどに遡って両者の交流がどれほどだったか推定したとしても、直近35年には遠く及ばない規模だったであろう。
ここ二千数百年の交流だけを通して、なぜO2b1aとO2b*が日本で29.9%、韓国で37.7%という規模で共有されているのか?なぜO2b1aとO2b*の比率が日韓で逆転しているのか?
なぜ朝鮮半島に見られるO1aとO2*またはO2a*が日本にまったく存在せず、ただO2a1*だけが認められるのか?なぜベトナムやジャワ島で一定数認められるのか?なぜミクロネシアではO2b*が数%の頻度で存在しているのか?そこになぜO2b1aはいないのか?
してみるとこういった事実は次のような考えに私たちを導いてゆく。
二千数百年前はおろかさらに時代を遡り、日本列島が大陸と地続きだった頃に発端を求めるのがふさわしいのではないか?最初に日本列島や朝鮮半島に定住した人々が、それらの遺伝子変異(SRY465/P49や47z)を共有していたのではないか?と。
大陸から日本列島が切り離された後で、片方でO2b1aがより多く増え拡がり、他方ではO2b*がより多数派になったと推定する。この方がより合理的だと思われる。
本稿の文献はこちらを参照されたい。
日本人のルーツ(Ⅷ)日本人はどこから来たのか?
2014.1.26
日本人はどこから来たか? 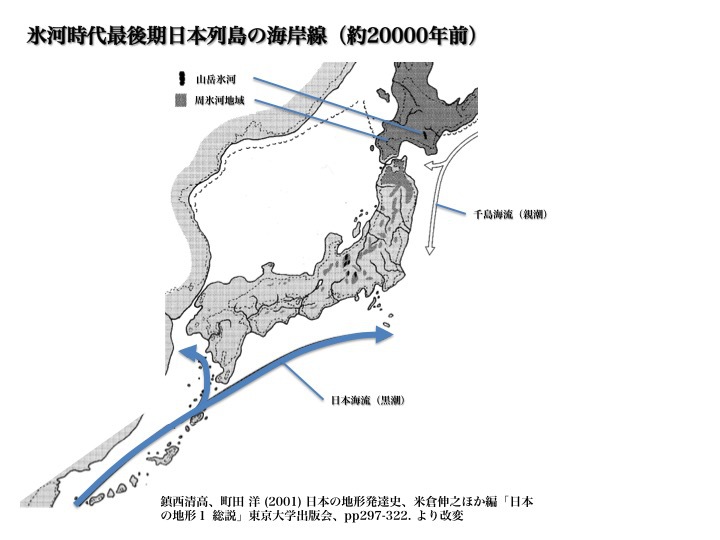
日本が大陸と地続きだった頃
本稿の文献はこちらを参照されたい。
日本人はどこから来たか?
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
このページのまとめ
1)旧石器時代後期の太平洋沿岸部の地形は現在と違う様相を呈していた。
2)日本が大陸と、北米大陸がシベリアと繋がっていた。
3)東南アジアにはスンダランド亜大陸が存在した。
4)東アジアへの移動は、南方の海岸線沿いルートと中央アジアルートが想定されていた。
5)常染色体ハプログループの解析は、西→東ではなく、南→北の移動を支持した。
6)Oのサブハプログループを再検討すると、海岸線沿いルートでも矛盾はなかった。
7)どちらも南方の海岸線沿いルートで、CとDの第一波が日本にたどり着き、
8)次にOの中のO2b、O2b1a、O3a2、O3a2c1が日本に到来した。
ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー
Ⅰ)これまで常識とされてきた概念との比較
前項で明らかになった現代日本人Y染色体DNAハプログループの特徴は、これまで常識とされてきた日本への人の流れに関する概念と、どのように整合性を保てるだろうか?
科学雑誌として一定の評価を得ている「ニュートン」2009年に載った図を見てみよう。
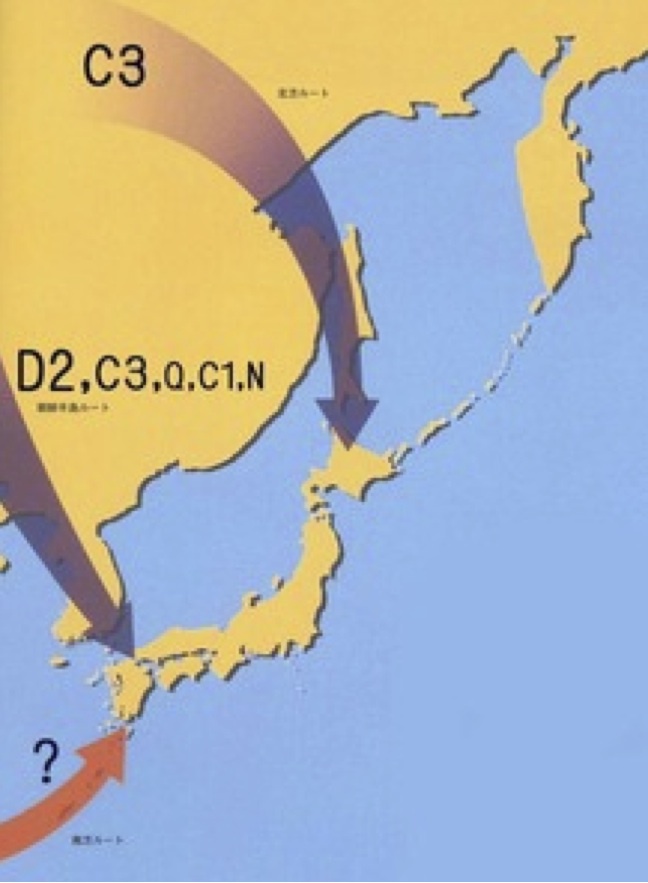
YdnaハプログループC3/D2/O/C1/Nの想定日本到達経路(左)
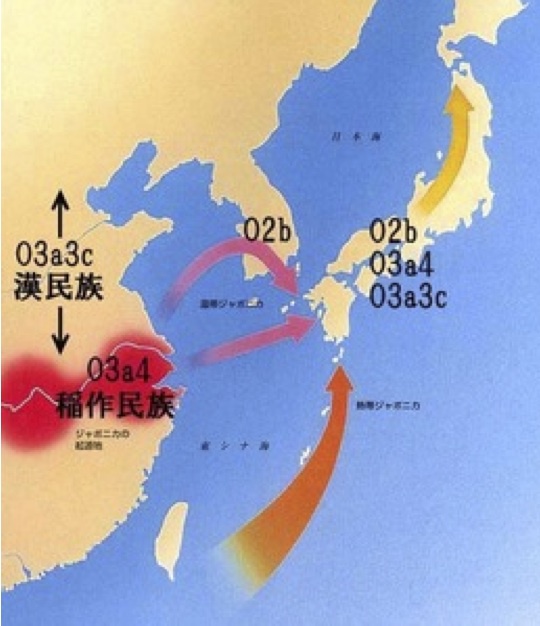
稲作文化をもたらしたと信じられていたYdnaサブハプログループO3a4(右)
<ニュートン・ムック>「最新・日本人の起源」ニュートンプレス、2009年による。
当時の考えを反映して、左図には樺太から北海道に北方ルートで入ったC3、朝鮮半島経由で入ったD2、C3、O、C1、Nが描かれ、南方ルートには「?」が付けられている。
右図には稲作文化の流入経路が描かれている。熱帯ジャポニカ米が南方ルートで、温帯ジャポニカ米が朝鮮半島経由または長江下流域から直接九州にたどり着いたと推定されていた。
温帯ジャポニカ米の水田稲作文化を携えて、O2b、O3a4、O3a3c集団が日本に入った。それが弥生人であるという想像である。
前述の通り、そもそもO2b集団に関しては、東南アジアに端を発するO2b*とO2b1aが長い旅をして、稲作文化が到来するはるか以前の日本列島と朝鮮半島近辺にたどり着き、一方は日本列島を中心に他方は朝鮮半島を中心に増え拡がった。それが合理的な説明と考えられる。
YdnaハプログループO3a4は2009年の古い分類による名称で、最新の国際遺伝系図学会のデータ(ISOGG 2013)ではO3a1cと変更されている。また同様にO3a3cはO3a2c1と名称変更されている。
既述の通り、現代人のハプログループの分布域は詳細に調べられている。O3a1c(図のO3a4)は華南、華北に分布するが日本列島にはわずかしかいない(1.1%、Nonaka 2007)。
一方、O3a2c1集団(図のO3a3c)の主な分布は華北(34.1%、Yan 2012)、華南(21.5%、Yan 2012)、チベット(34.3%、Hammer 2006)、東南アジア(4.2~27.9%、Hammer 2006)で、日本列島にも7.6%存在する(Nonaka 2007)。
華南の長江文明集団を形づくっていたのは、O3a2b(モン族、ミェン族)、O1a、O2a1とされている。しかしそれらの亜型は、日本にほとんどわずかしか、あるいはまったく存在しない。
これは一体何を意味するのだろうか?長江文明集団の日本列島への直接関与は少なかったということである。
ただ日本列島に一定数存在するO3a2c1(図のO3a3c:約10.5%または7.6%、Hammer 2006, Nonaka 2007による)は、長江文明にどれほど関与した人たちだったのだろうか。
日本で以前O3として一括りにいわゆる弥生系とされていたグループの大部分は次の通り、華南では多数派とはいえないO3a2と、華南のみならず華北、チベット、東南アジアに拡がるO3a2c1集団の二つのサブハプログループ(日本人全体の20%前後(Nonakaのデータでは約12%))である。
日本列島にしかいないC1、D2、ほぼ日本列島のみのO2b1a、シナ大陸にはあまりいないが日本で見つかっているO2b*とO3a2、それにC3をあわせて、86.2%(81.8%)にのぼる。日本人に見つかっていてシナ大陸にも存在するハプログループは12.3%(13.7%)にすぎない。
そのうち10.5%(7.6%)を占めるO3a2c1集団が日本に来た渡来系弥生人だろうか。他のグループ、特にO3a1cやO2aはどうしたのだろうか。O1aもNonaka 2007のデータでは見つかっているがHammer 2006のデータでは認められない(下線はHammer 2006のデータ、括弧内下線はNonaka 2007のデータによる)。
彼らO3a1c、O1a、O2a、O3a2b集団、つまりO3a2c1以外の人々が来なかった理由、来たけれども増えなかった理由、または絶えてしまった理由を考えなくてはいけない。また華南の少数派O3a2が、現代日本で何故4.2%もの頻度を占めているのか、その説明も必要である。
つまり、稲作技術を持ったボートピープルがシナ大陸から少数やって来て、O3a2c1を持つグループの人口だけが弥生人として爆発的に増加していったという可能性を論じることになる。このO3a2c1集団人口爆発説は妥当な仮説だろうか。
爆発的な増加を、O3a2c1集団が先住縄文人を征服した結果と見る人もいるだろう。もしそうであるなら、征服民である彼らは何故彼らの言語を被征服民に使わせなかったのだろうか。今のところ、日本語の骨格や発音に影響を及ぼした痕跡はほとんどなく、借用語としていくつかの例が示されているにすぎない。
そもそもボートピープルがやってきた規模は、「押し寄せてきた」とは程遠く、「年に二~三家族、パラパラとやって来た」に過ぎなかったという(「日本人のはるかな旅」5)。いわゆる縄文人の村の遺跡はたくさんあるものの、渡来人だけのムラの遺跡はこれまで発見されていない。
こうしてみると、渡来系弥生人が日本人のルーツであるという説は根拠が非常に薄いようである。日本と朝鮮半島、日本とシナ大陸という二地域関係だけに焦点を絞らず、よりマクロな視点に立ち、もっと合理的な説明があればそれを採用すべきだという意見に賛成する。
Ⅱ)アダムの息子たちの旅の背景
そこで少し視点を変えて拡げ、人類がアフリカを出て長い旅をし、日本にたどり着いて住み着いた頃の背景を見てみることにする。
ⅰ)68000-30000年前の世界
約68000-60000年前にC、D集団が出アフリカを果たした頃の地球はどのような環境だったのだろう。
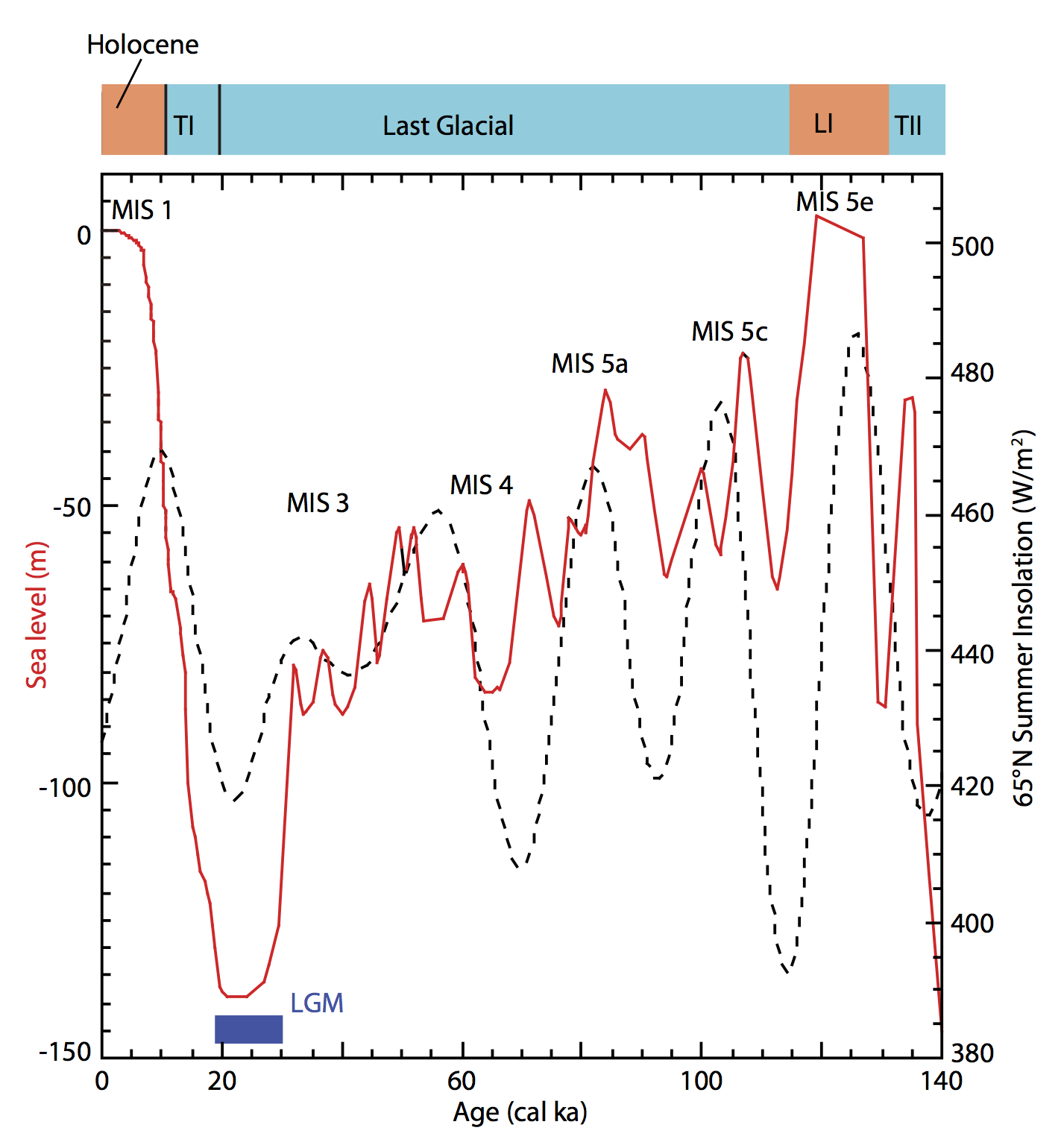
約140000年前から現在までの海面レベル(赤実線)
横軸に年代をとり(単位は千年、右端が140000年前、左端が現在)、縦軸左に現在と比べた海水準(m)をとっている。
Holoceneは完新世で、地質学の第四紀最後期、つまり現在に続く最も新しい区分を表す。
Last Glacialは最終氷期、TI、TIIは氷期の終末期、LIはLast Interglacial最終間氷期を示す。
LGMはLast Glacial Maximum最終氷期極期である。
MISはMarine Isotope Stage海洋酸素同位体ステージで、酸素安定同位体18Oの比による過去の気温を推定する方法を根拠にした時代区分。海底堆積物をボーリングし、深海底の有孔虫の殻の酸素同位体比率から、海洋水温の変動を推測し再構成する。最終氷期とその前後は、MIS 1~MIS 5cまで区分されている。酸素安定同位体18Oの比による過去の気温を推定する原理の詳細はδ18Oの項(後述)を参照されたい。
縦軸右と黒い破線は北緯65度で日光がどれくらい当たっていたかであるが、本稿では割愛する。 Yokoyama et al. 2011 Oceanography による。
この図からもわかるように、当時は氷河期だった。ほとんどは最終間氷期(LI)と最終氷期後の現在に挟まれている、最終氷期(最新氷期)と呼ばれる時代だった。
図では約140000年前から現在までの推定海面レベルが赤の実線で示されている。海水準は最終氷期極期(LGM=Last Glacial Maximum)に-140m近くに達し、50000-30000年前には-85~-55m程度を推移したとしている。
新人が出アフリカを果たした約68000年前頃の海面レベルはおよそ-60~-75mだった。アフリカの角と呼ばれるソマリアの隣国ジブチとアラビア半島南端のイエメンを隔てるバブ-エル-マンデブ海峡は現在よりも狭く、工夫すれば少数の人々が渡れるほどだった(次図参照)。
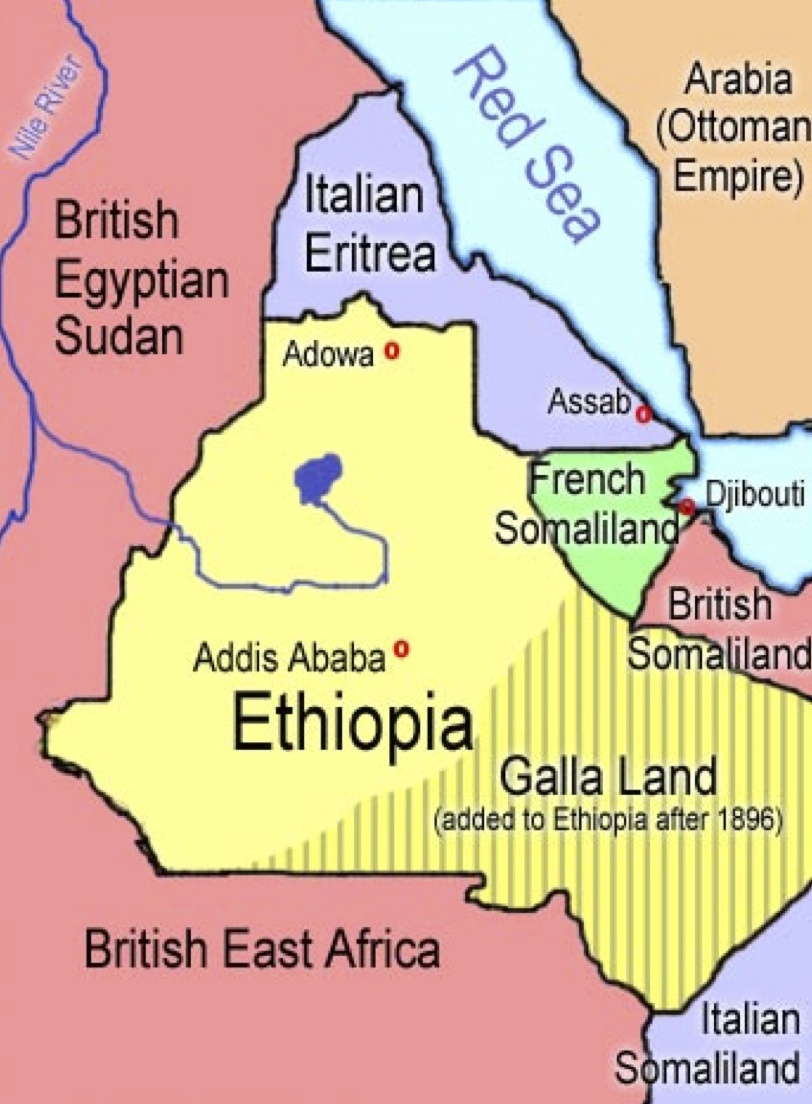
植民地時代のエチオピア周辺
フランス領ソマリランド(French Somaliland:緑)が現在のジブチ共和国である。
British Somaliland:英領ソマリランド、
Italian Somaliland:伊領ソマリランド、Italian Eritrea:伊領エリトリア、
Ethiopia:エチオピア、British Egyptian Sudan:英領スーダン、
British East Africa:英領東アフリカ、
Arabia (Ottoman Empire):アラビア(オスマン帝国)。
Wikipediaによる(CC BY-SA 3.0)
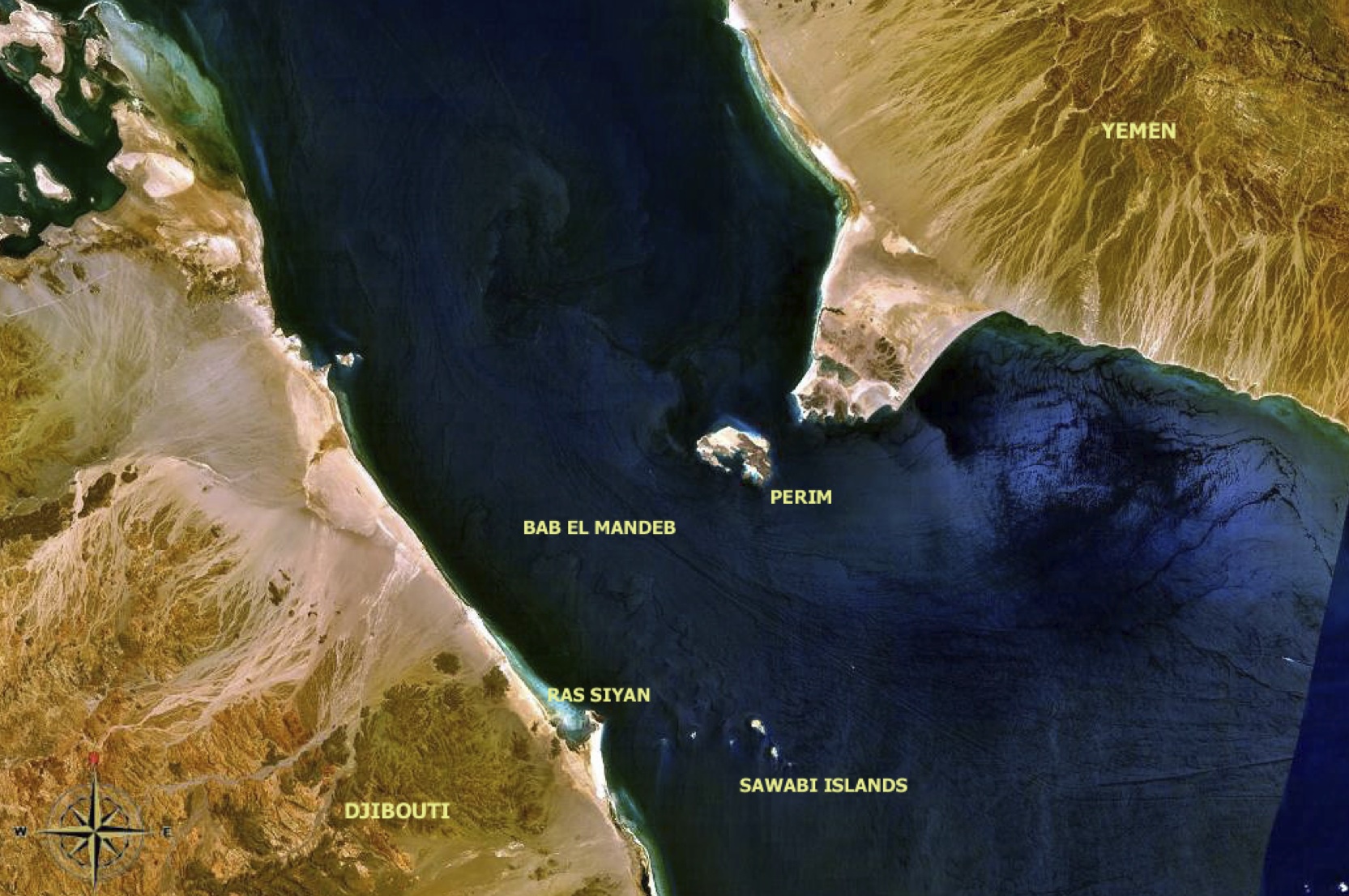
バブ-エル-マンデブ海峡
左が現在のジブチ共和国、右がイエメン共和国である。幅は30kmほど。だが最終氷期時代には11km程度に縮まっていたと考えられる。
写真はNASAによる。
イエメンを経て反時計回りで海岸沿いを行くとペルシャ湾に至ることができる。現在平均水深50mほどのペルシャ湾は当時周囲から河川が流れ込む緑豊かな渓谷だったとされる。エデンの園に比定する研究者も存在するほどだという(次図参照)。

アラビア半島とイランに囲まれたペルシャ湾
半島海外沿いを反時計回りに進むとペルシャ湾に到達した。
最終氷期時代には湾のかなりの部分が海水準より高く、
豊かな渓谷だったと推定されている。
写真はNASAによる。
約50000-40000年前の時期においては約-80m程度海水準が低下していた。当時の東南アジア、日本列島とその近辺は次の二つの図のような地形をしていたと推定される。
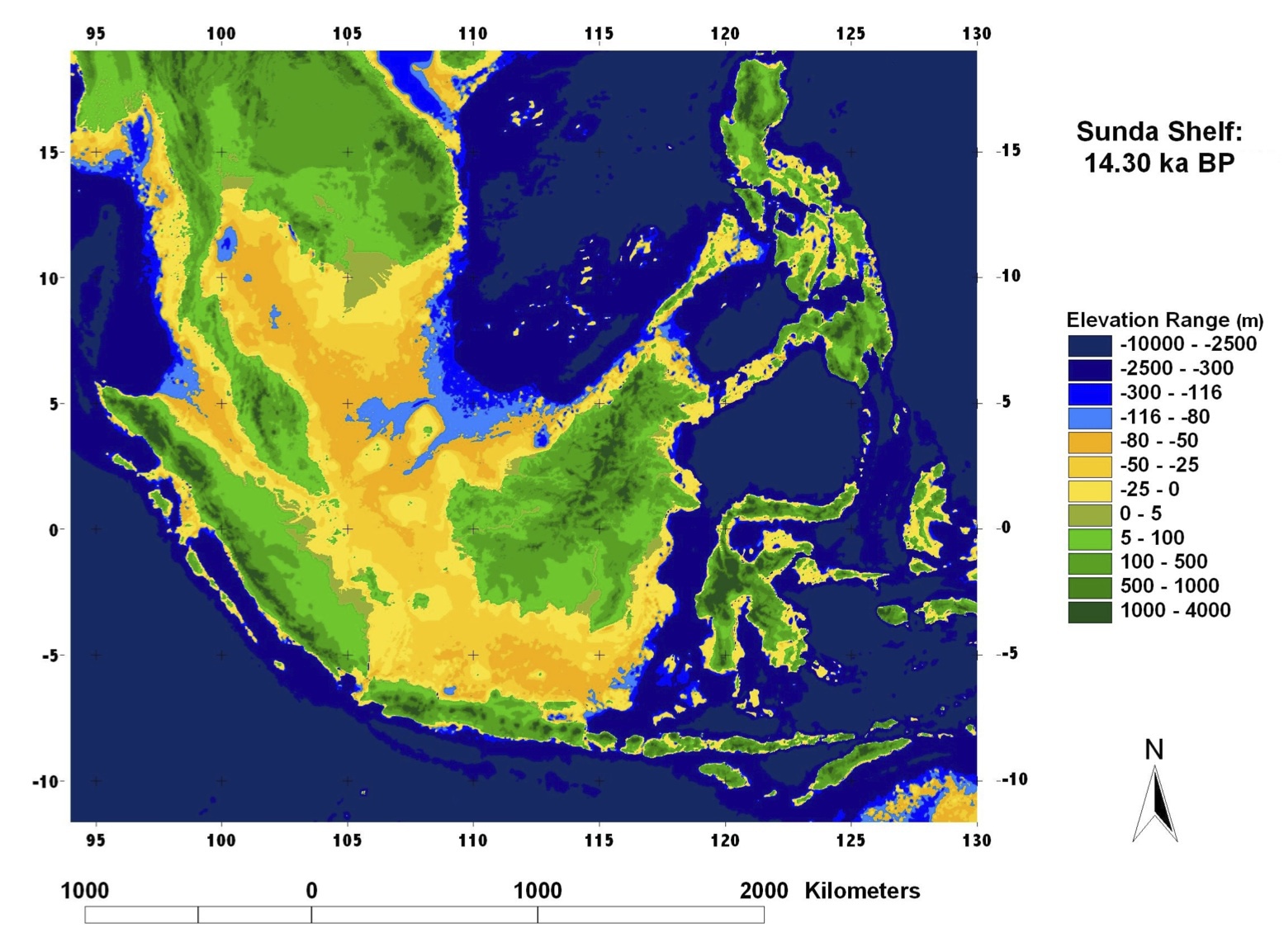
海面レベル-80mの東南アジア
Voris 2000より改変。
http://www.fieldmuseum.org/explore/pleistocene-sea-level-maps
東南アジアには広大な陸地が存在しておりスンダランド(スンダ亜大陸)と呼ばれていた。アジアの南海岸沿いを旅して来たアダムの子孫たちは、温暖なスンダランドで狩猟採集生活を送ったと考えられる。
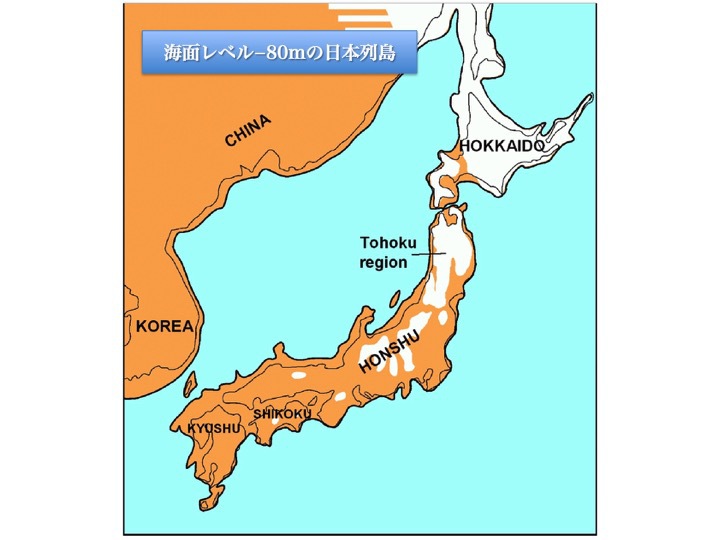
海面レベル-80mの日本列島
Wikipediaによる(CC BY-SA 3.0)
日本列島はといえば、現在の間宮海峡の最浅部は8m、宗谷海峡の最浅部は30-70mで、どちらも大陸と地続きだったと考えてもよい。他方、津軽海峡の最浅部は140m、対馬海峡の最浅部は130m程度で、どちらも眼前には海が広がっていた。
海水準-80mといえど、直線距離で40km近くもあった対馬海峡西水道(朝鮮海峡)を、人々が渡ることは困難だったと思われる。ただ津軽海峡は冬期間中に海が凍っていれば渡海が可能だっただろう。
ⅱ)30000-20000年前の世界
約30000-20000年前の地球はどのような環境だったろう。最終氷期極期で、後期旧石器時代に相当する。
まず先史時代の地球の気候を解く一つのカギについて説明しよう。δ18Oである。

δ18Oとは?
このδ18Oとは上図の数式で計算される酸素同位体の割合(‰)のことである。
グリーンランドは厚い氷で被われている。氷床と呼ばれる。氷床は降り積もった雪が固まり圧縮されてできた。年輪のように何層にも何層にも重なっている。
気象学者は18Oという安定同位体に着目した。水分子は16Oの通常の酸素から成り立つものがほとんである。だが一定の割合で18Oという安定同位体の酸素から成り立つ水分子も存在する。グリーンランド氷床の氷を調べ、18Oと16Oの割合が海水温の影響を受けることを見出したのだ。
原理はこうだ。16Oを含む軽い水ほど先に蒸発し雲となる。その雲から雪が降り長年蓄積して氷床を形成する。降雪年の平均海水温度が低いほど、氷床氷サンプルにおける18Oの割合は小さくなる。つまりδ18O値が小さいほど低海水温であったことを示すわけである。
こうして測定したδ18Oから当時の海水温、すなわち地球の外気温が推定される。次の図の上のグラフは、δ18Oを縦軸に年代を横軸にとってあり、当時の気温変動の推移が示されている。
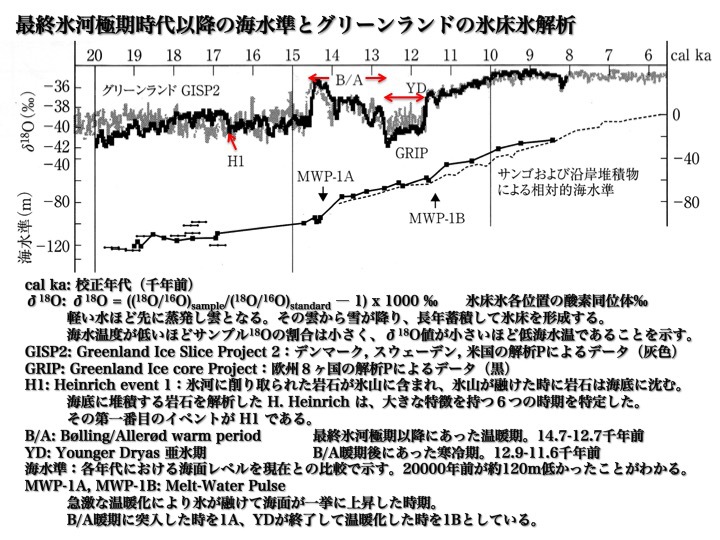
最終氷河極期以降の海水準とグリーンランド氷床氷解析
太田陽子、小池一之、鎮西清高、野上道男、町田 洋、松田時彦著「日本列島の地形学」2010、p108より。
cal ka: 校正年代(千年前)。左端が約20000年前、右端が現在を示す。
δ18O: 氷床氷各位置の酸素同位体の割合い(‰)。
GISP2: Greenland Ice Slice Project 2:デンマーク, スウェーデン, 米国の解析プロジェクトによるデータ(灰色)、
GRIP: Greenland Ice core Project:欧州8ヶ国の解析プロジェクトによるデータ(黒)。
H1: Heinrich event 1(ハインリッヒ事象):氷河に削り取られた岩石が氷山に含まれ、氷山が融けた時に岩石は海底に沈む。海底に堆積する岩石を解析した H. Heinrich は、大きな特徴を持つ6つの出来ごとを特定した。その第一番目が H1 である。
B/A: Bølling/Allerød warm period(ボーレン/アレレード温暖期)、最終氷河極期以降にあった温暖期。14.7-12.7千年前。
YD: Younger Dryas(ヤンガー・ドライアス)亜氷期、B/A温暖期後にあった寒冷期。12.9-11.6千年前。
海水準:各年代における海面レベルを現在との比較で示す。
MWP-1A, MWP-1B: Melt-Water Pulse(融解水衝撃)、急激な温暖化により氷が融けて海面が一挙に上昇した出来ごと。B/A暖期に突入した時期を1A、YD亜氷期が終了して温暖化した時期を1Bとしている。
この図の下のグラフでは、後期旧石器時代における推定海面レベルが示されている。約20000年前の推定海水準は約ー120mだった。6000年かけてゆっくりと海岸線は後退する。
約15000年前から急激に300年間で20mというペースで海面が上昇する。気温が一気に上昇して多くの氷が融けたために、その衝撃が世界に及び海水準が急に上がったのである。これが融解水衝撃(MWP-1A)と呼ばれるものだ。
次の2000年は約25mの海面レベル上昇とその速度がゆるやかになる。だが11500年前に海水準が急に上がる時期がくる。これが二度目の融解水衝撃(MWP-1B)と称される。その後やがて現代の海岸線にほぼ一致するところにまでなる。
こうしてみると、最終氷期極期の特徴として、海面レベルが現在よりはるかに下がっていたことが挙げられる。次図は後期旧石器時代の太平洋沿岸部の地形を示すもので、特に緑で囲ったような海岸線となっていたと推定される。
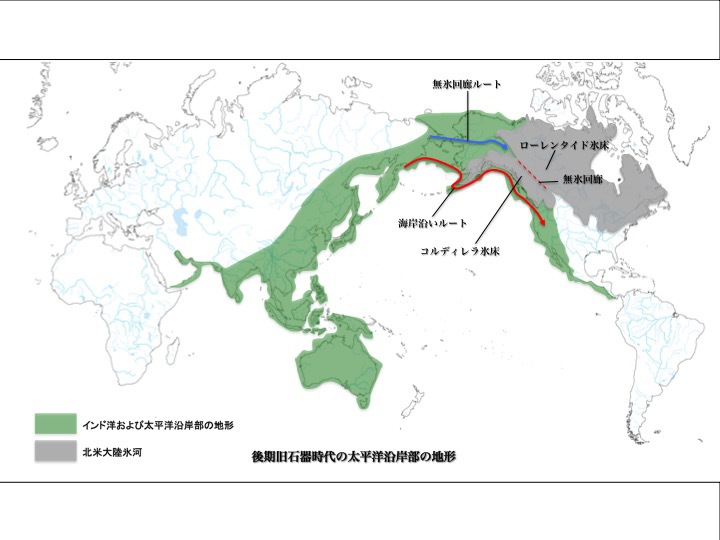
後期旧石器時代の太平洋沿岸部地形
松本克己「世界言語の中の日本語」より改変。
最終氷期極期すなわち後期旧石器時代、東南アジアにはスンダランド大陸が、その南東にはサフールランド大陸が存在していた。次図に示すとおりである。
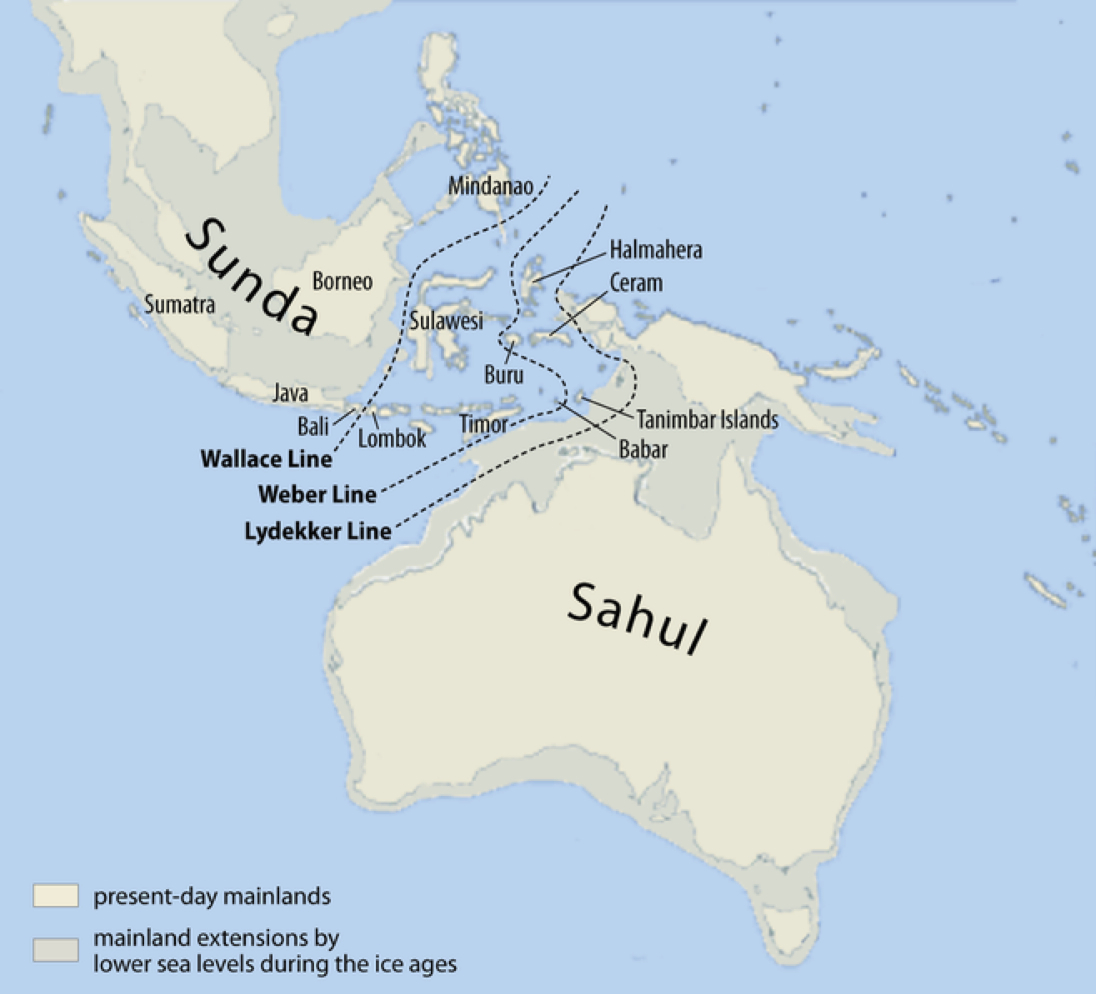
スンダランド(Sunda)とサフールランド(Sahul)
Sumatra:スマトラ島、Java:ジャワ島、Bali:バリ島、Lombok:ロンボク島、Timor:ティモール、Borneo:ボルネオ島(カリマンタン島)、Sulawesi:セレベス島(スラウェシ島)、Mindanao:ミンダナオ島、Halmahera:ハルマヘラ島、Buru:ブル島、Ceram:セラム島、Tanimbar Islands:タニンバル諸島、Babarバーバル島。
Wallace Line:ウォレス線、Weber Line:ウェーバー線、Lydekker Line:ライデッカー線。これらの線を挟んで動植物の生態系が異なっていることが知られている。
Wikipediaによる(CC BY-SA 3.0)
次の衛星写真からも当時陸地だった部分が薄水色で容易に想像可能である。
ランドサット衛星の写真を合成した東アジア
GoogleEarthより
次図は約21000年前のスンダランドの推定海岸線と地形である。
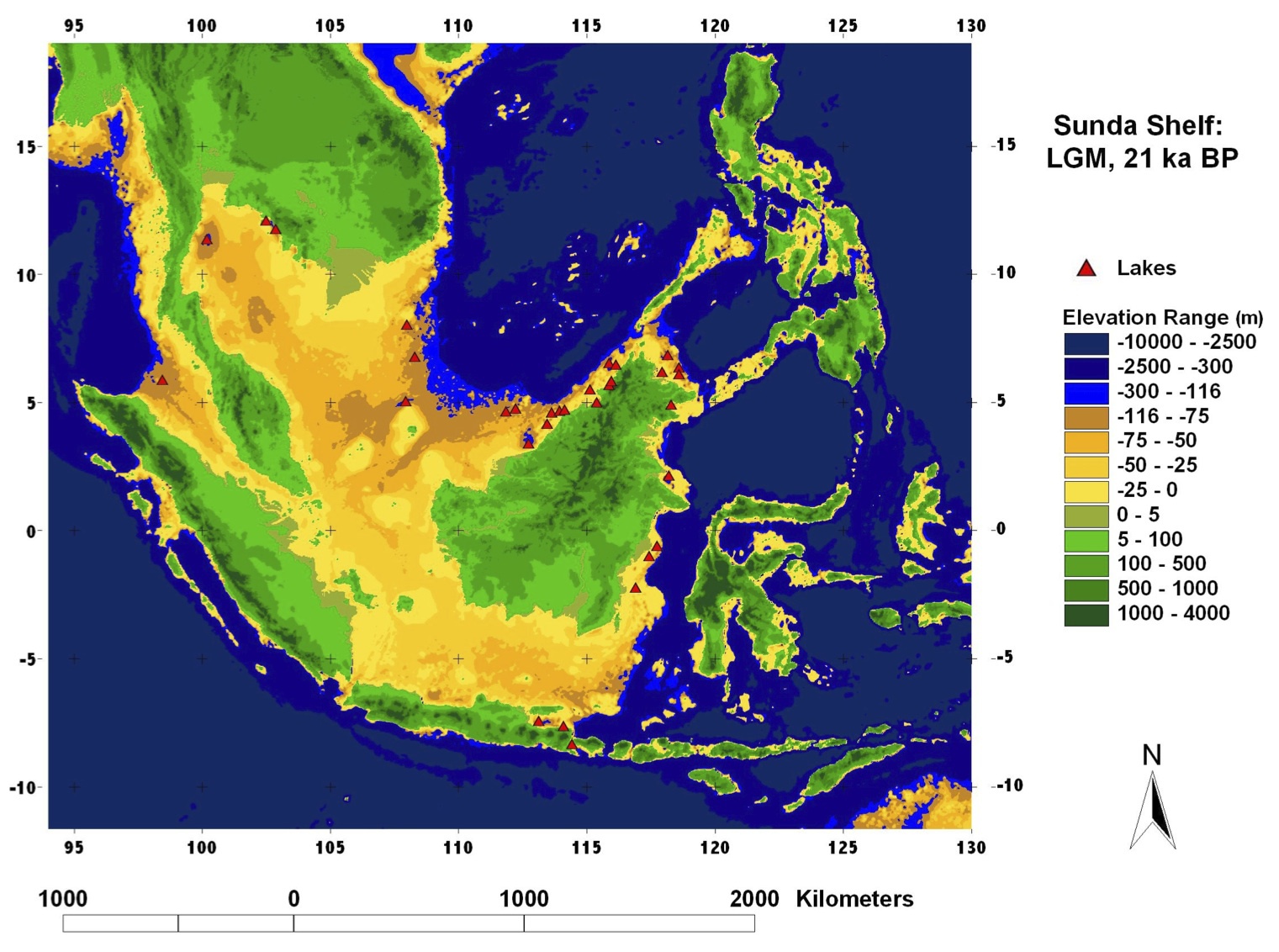
約21000年前のスンダランド推定地形
海面を現在比ー116mと推定してイラスト化している。
Voris 2000より改変。
http://www.fieldmuseum.org/explore/pleistocene-sea-level-maps
スンダランドの消失シミュレーションがカラーのイラストとして載っている。
こちらを参照されたい。
当時の推定海面レベルをプロットしてグラフ化したのが次図である。約15000年前のMWP-1Aによるが急激な海水準上昇が特徴的に見て取れる。
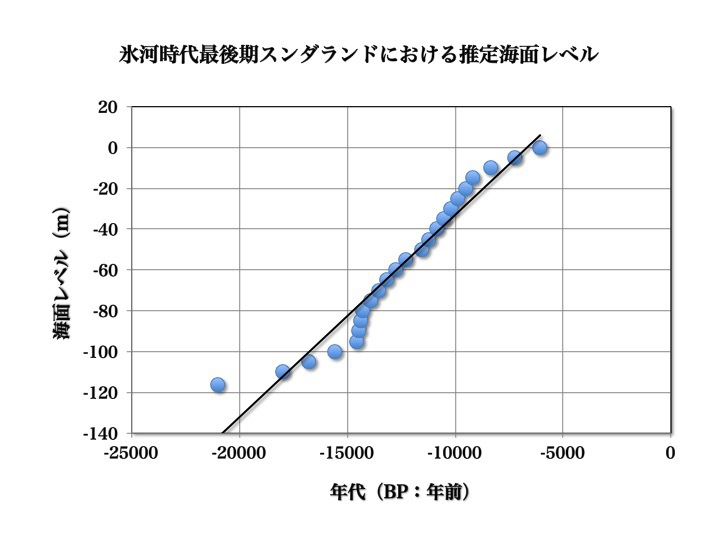
氷河時代最後期における推定海面レベル
縦軸に現在と比べた推定海面レベルをとり、横軸に時間をとってプロットしたもの。
Voris 2000より改変。
ⅲ)最終氷河期極期の日本列島
では最終氷期極期(LGM)だった頃、日本列島はどのような姿だったのだろう。おそらく次の図が最も参考になるだろう。
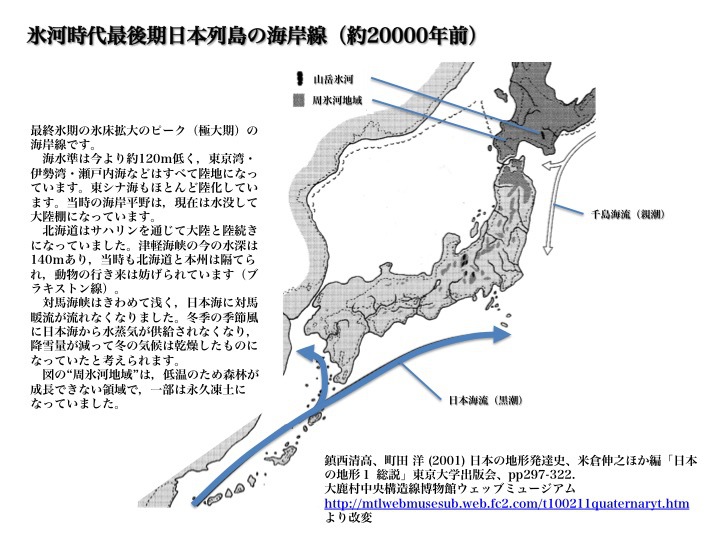
最終氷期最後期(後期旧石器時代)日本列島附近の海岸線
鎮西清高、町田 洋 (2001) 日本の地形発達史、米倉伸之ほか編「日本
の地形1 総説」東京大学出版会、pp297-322、
および
大鹿村中央構造線博物館ウェッブミュージアム
http://mtlwebmusesub.web.fc2.com/t100211quaternaryt.htm
より改変。
これは約20000年前、最終氷期極期、後期旧石器時代の日本列島付近の海岸線を示している。海岸線レベルは今より約120m低く、東京湾、伊勢湾、瀬戸内海などはすべて陸地になり、東シナ海もほとんど陸化している。
北海道はサハリンを通じて大陸と陸続きになっていた。津軽海峡の現在の水深は140mで、当時も北海道と本州は隔てられ、動物の行き来は妨げられていた(ブラキストン線)。図の“周氷河地域”は、低温のため森林が成長できない領域で、一部は永久凍土になっていた。
対馬海峡はきわめて浅く、狭く、大きな川のような様相を呈していた。当時、日本海には対馬暖流が流れていない。冬季の季節風に日本海から水蒸気が供給されず、降雪量が減り冬は乾燥したものになっていた。
このような対馬海峡なら容易に渡れる。そこを通って人々が日本列島に流入したと考えるのは合理的である。
ちょうど最終氷期極期(LGM)の約30000-20000年前頃は、ユーラシア大陸と北米大陸を結ぶベーリング海も陸化していた。ハプログループC、Qの集団の一部がユーラシアから北米への大移動をしていた時期に一致している。
ⅳ)日本にたどり着いたO集団
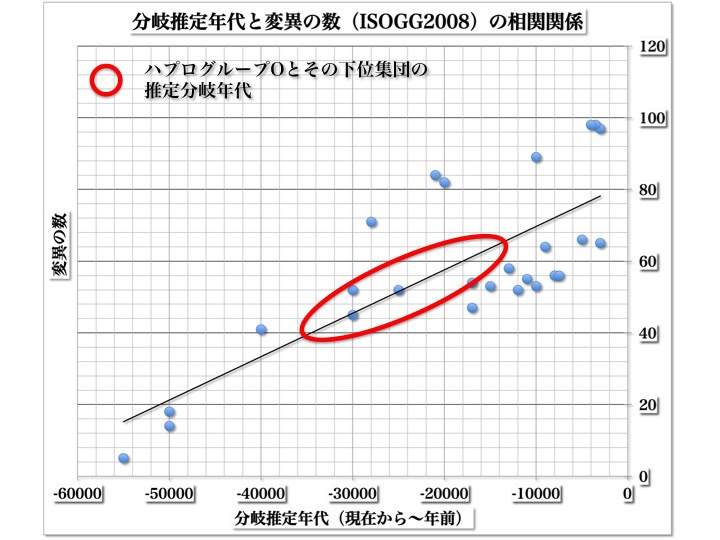
YdnaハプログループOとそのサブハプログループの推定分岐年代
縦軸に各ハプログループが分岐するまでに累積した変異の数をとっている。BTの最初の変異を1個目として計算した。横軸に時間をとった。各点はヨーロッパに分布するハプログループ、サブハプログループの推定分岐年代データと上述のような累積変異数をプロットしたもの。
http://en.wikipedia.org/wiki/Human_Y-chromosome_DNA_haplogroup にある、Chronological development of haplogroups in Europe にある各グループの推定分岐年代を用いた。
最小二乗法で直線を引き、ハプログループOとそのサブハプログループの累積変異数から推定される分岐年代を赤い円で囲んだ。
変異の数は、ヨーロッパに分布する各グループの分岐年代を推定した時期のデータに対応するよう、世界遺伝系図学会(ISOGG)の2008年の系統樹によった。
日本にたどり着いたYdnaハプログループC、D、Oのうち、O集団がそのグループ内で分岐していった時期はいつ頃なのだろうか。上の図からは、Oとその下位集団の推定発生および分岐完了年代が、約35000年から15000年前と計算される。
ちょうど最終氷期極期が始まる頃から、MWP-1A(融解水衝撃)による急激な海水準上昇直前までの時期に重なっている。
ⅴ)東南アジアへの人類移動定住のモデル
アフリカを出発した人々がアジアに来たのはいつ頃のことか。インドネシアを例にとって、最近の説を紹介する。
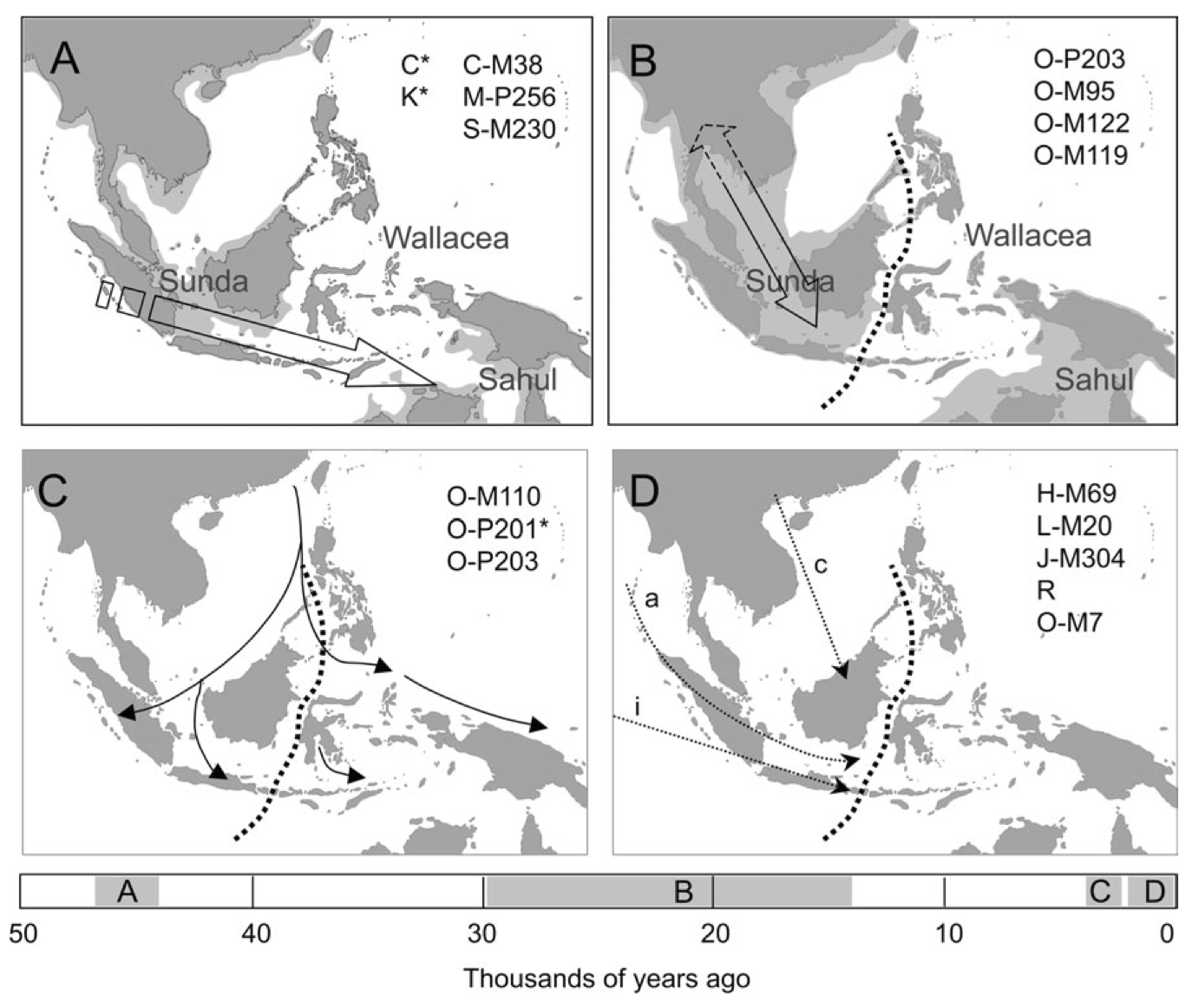
インドネシア列島への人々の到達と定住のモデル
Karafet 2010による。
図上の四つは(A)50000-40000年前、(B)旧石器時代の人々の流れ、(C)オーストロネシア系集団が稲作農業を携えて海洋進出していった頃、(D)歴史時代におけるヒンドゥー教の波及(3~13世紀)、アラビアからイスラム教が波及した時代の人々のインドネシア列島への流入が描かれている。
図(A)では海面レベルー50m、(B)では海面レベルー120mとして、該当する年代の海岸線と陸地を薄いグレーで示している。
図下は、時間軸をとって(A)~(D)それぞれの集団の波が列島に来た時期を推定している。
図(A)のC-M38、M-P256、S-M230はそれぞれハプログループC2、M、Sを特徴付ける変異名、
図(B)のO-P203、O-M95、O-M122、O-M199はそれぞれハプログループO1a1、O2a1、O3、O1aを特徴付ける変異名、
図(C)のO-M110、O-P201*、O-P203はそれぞれハプログループO1a2、O3a2*、O1a1を特徴付ける変異名、
図(D)のH-M69、L-M20、J-M304、O-M7はそれぞれハプログループH、L、J、O3a2bを特徴付ける変異名である。
この図の下の時間軸にグレーで示された到達時期に注目する。このモデルによると、約50000-40000年前に最初の集団(C、M、S)がやって来た。
約30000-15000年前、後期旧石器時代に次の集団(O1a、O1a1、O2a1、O3)が到来した。さらにオーストロネシア系集団(O1a1、O1a2、O3a*)の海洋進出が約3000-2000年前と推定されている。
ⅵ)日本列島へのC、D、O集団の到来
日本列島でも上記のモデルと似たような傾向を示すと仮定するとどうだろう。すなわちDとC集団が約30000年よりも前の時期に日本に第一陣として到達し一部が定住。次いで約30000-20000年前頃に、O2b1a、O2b、O3a2、O3a2c1など第二陣がやって来たとするのだ。
約30000年よりも前の時期だとすると、北の樺太、北海道経由で、冬期氷結した津軽海峡を渡って来たという説が有力になる。第二陣は最終氷期極期の約30000-20000年前になる。
第一陣がもっと遅く最終氷期極期の前半(約30000-20000年前)に来たと仮定すると、ほとんどは大陸と地続きだった対馬経由と考えられる。第二陣は最終氷河期極期の後半(約20000年前頃)から急激な海面レベル上昇直前(約15000年前頃)までというのが妥当な線だろう。
もちろん第一陣の一部が北方から日本列島に到来したことを完全に否定するわけではない。
第二陣として日本列島に到達したと目されるハプログループO集団は、最終氷期極期にはさかんに移動していた。前項で一つのモデルを紹介したように、インドネシアを含む他の地域には展開していたが、日本にだけは来なかったとする方が奇妙な話だろう。
いずれにせよ、第二陣でもって原日本人の日本列島への移住がひと段落しただろう。列島は大陸とほとんど地続きだった。最終氷期極期、後期旧石器時代の頃である。
その後、最終氷期の終焉とともに海岸線がどんどん上がる。特に約15000年前の急激な海水準上昇により(MWP-1Aが引き金となった)、日本列島は大陸と切り離され、人々の往来が困難となったからである。
前述したように、ちょうど約30000-20000年前、最終氷期極期(LGM)の頃は、陸化していたベーリング海を経由してハプログループC、Qの集団がユーラシアから北米への大移動をしていた時期である。
ハプログループC、D集団が日本に流入した時期も、ちょうど同じような約30000-20000年前頃と考えるのが妥当なところだろう。
以上から、本稿では、日本人に特徴的なC、D、Oの各集団は、今から約30000~15000年前頃に日本列島に入って来たという立場をとることにする。第一陣はCとD、第二陣がO集団である。それが最も合理的な説明だと思われるからである。
Ⅲ)東南アジアから東アジアへの移動
ⅰ)南方ルートvs中央アジアルート
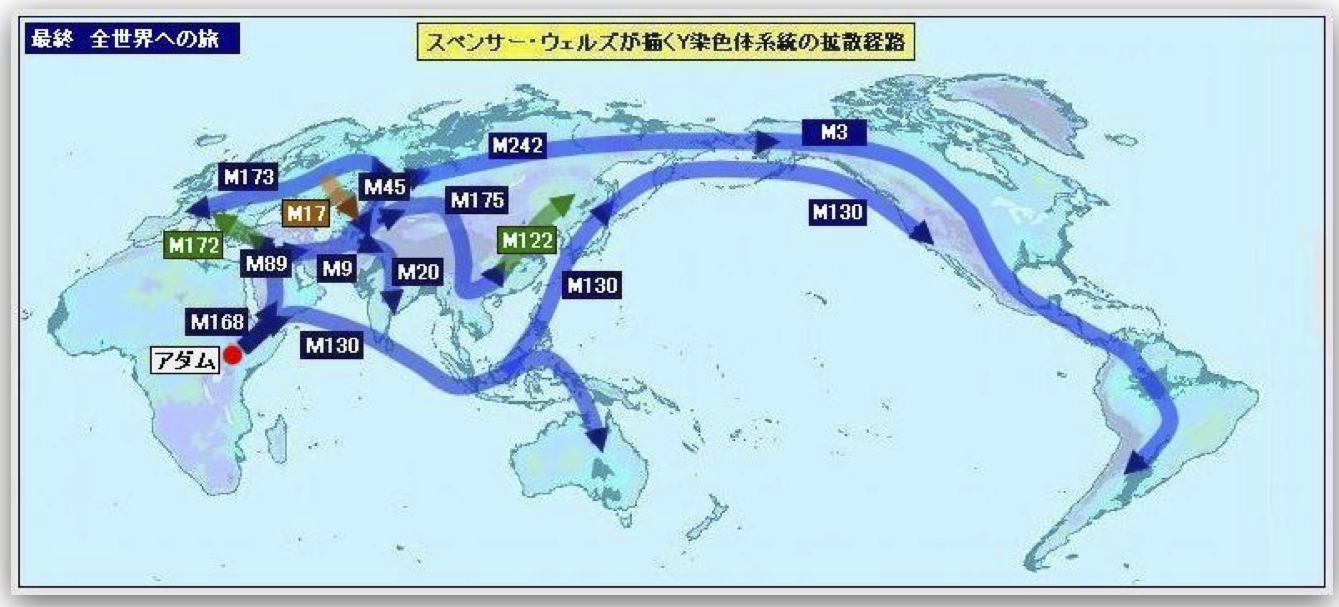
スペンサー・ウェルズの描くYdnaの世界への拡散経路
http://www.geocities.jp/ikoh12/kennkyuuno_to/012_3adam_no_tabi_syouhonn.html
(伊藤俊幸氏による図)より
M172、M168などは変異の呼称、M130はハプログループC、M175はハプログループO、M122はサブハプログループO3を特徴づける変異である。
ハプログループC(M130)は海岸線沿いに、アラビア半島南端部、ペルシャとインドの海岸部を経て東南アジアに渡り、そこからオーストラリアに行ったり、北上して日本列島に経由してシベリア、カムチャッカ、アリューシャン列島、北米大陸西海岸を南下したり大移動をした。
スペンサー・ウェルズはYdnaハプログループ解析から、一人のアダムから子孫がどのような経路で世界に移動拡散していったかを推定した。特に中央アジアの重要性について論じた(Wells 2001)。
ハプログループOは、その祖先F(M89)、K(M9)から中央アジアの草原で枝分かれした(M175)。その後東南アジアに南下し、またシナ大陸を北上してサブハプログループO3(M122)に分岐したと考えた。
上述のCと同様に、ハプログループDも海岸線沿いに南回りで東南アジアを経由し、日本にたどり着いたと考えられている。C、D以外、つまりハプログループOに関してはどうなのだろう。
後期旧石器時代の人々の流れについては仮説が二つある。
(A)ひとつ目は、すべての東南アジア、東アジアの集団が南方の主に海岸線沿いの経路をたどってシナ大陸や日本を含む極東地域に入ったというもの。
(B)ふたつ目は、少なくとも二つのルートを通ったというもので、最初は南方ルート、後になってから、より北のルートを何度も通っていくタイプの移動を想定する。後者ではヨーロッパと東アジアの集団を橋渡しする形となり、東南アジアの貢献はほとんどないとしている。Wellsは後者の立場である。
ⅱ)南方ルートを支持する常染色体SNPハプロタイプの大規模データ解析
そこに、常染色体一塩基多型(SNP)の組合わせ(ハプロタイプ)を大規模に解析した研究が一石を投じた。サイエンス誌(The HUGO Pan-Asian SNP Consortium 2009)に掲載された論文のエッセンスを紹介する。
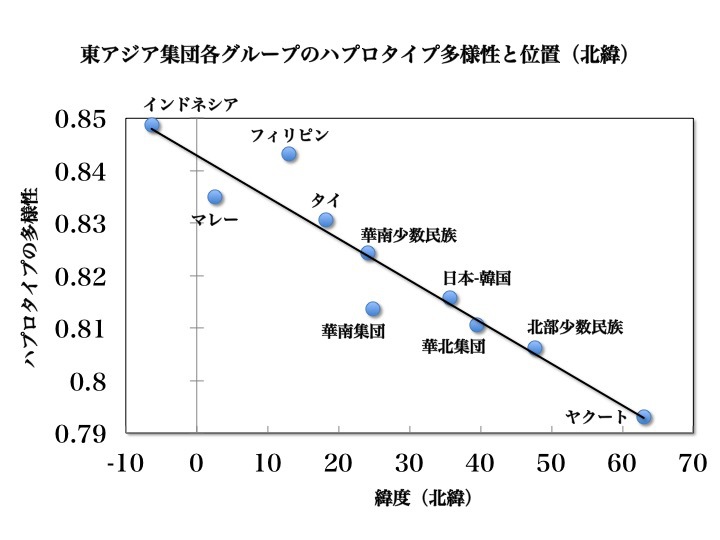
東アジア集団各グループにおける常染色体SNPハプロタイプの多様性と
各グループが暮らす緯度の関係
現代アジア人の常染色体SNP(一塩基多型)を大規模解析し、東アジア各集団が有するSNPハプロタイプの多様性と各グループが住んでいる位置(緯度=北緯)の関係をプロットしたデータ。縦軸はハプロタイプの多様性、横軸は北緯を表している。インドネシア人、マレー人、フィリピン人、タイ人、中華人民共和国(PRC)南方系少数民族、華南集団、日本人-韓国人、華北集団、PRC北方系少数民族、ヤクート族の位置を示してある。
The HUGO Pan-Asian SNP Consortium 2009 より改変。
上図は常染色体SNPハプロタイプの多様性と緯度の関係を解析したものである。SNPハプロタイプの多様性が南ほど著しく、北に行くほど少なくなっていることがわかる。
緯度の低いインドネシア、マレー半島、フィリピン、タイなど東南アジアのSNPハプロタイプの方が、緯度の高い南シベリアのヤクートや日本、韓国、華北、北部少数民族よりも多様性に富んでいたのである。
中央アジアルートなどを想定し、東南アジアは通らなかったという(B)が正しいなら、シナ大陸が最も多様性に富んでいて、東南アジアに下るにつれて多様性が薄らぎ、極東や北東アジアに行くにつれて多様性が減ってゆくことが予想される。
しかし上図のデータは東南アジアが最も多様性に富む地域であることを示しており、仮説(A)の方が正しいことを示唆している。
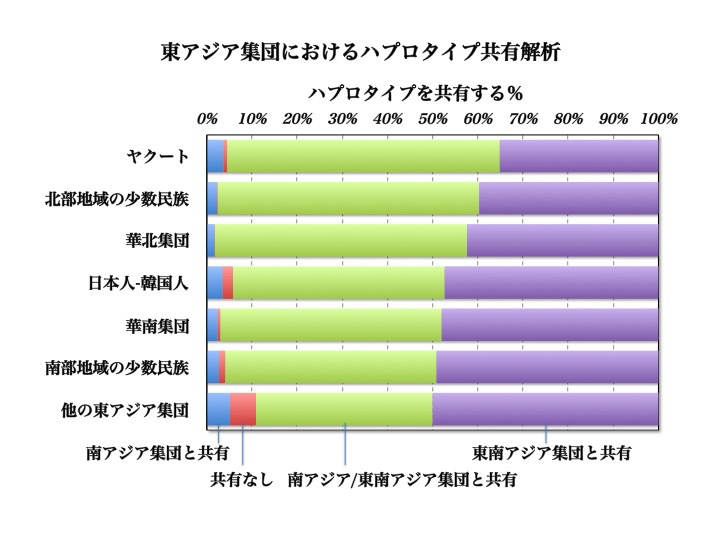
東アジア集団における常染色体SNPハプロタイプの共有解析
現代アジア人の常染色体SNP(一塩基多型)を大規模解析し、東アジア各集団が中央南アジア(CSA)および東南アジア(SEA)の集団と、どれほどSNPハプロタイプを共有しているかを%で表したデータ。縦軸には、上からヤクート族、中華人民共和国(PRC)北方系少数民族、華北集団、日本人-韓国人、華南集団、PRC南方系少数民族、他の東アジア集団全体、横軸にはハプロタイプを共有する%を示してある。青:南アジア集団とのみ共有する%、赤:南アジア、東南アジア集団いずれとも共有しない%、緑:南アジア、東南アジア集団どちらとも共有する%、紫:東南アジア集団とのみ共有する%。
The HUGO Pan-Asian SNP Consortium 2009 より改変。
この図は東アジア集団における常染色体SNPハプロタイプが、南アジア集団や東南アジア集団とどれほど共有されているかを解析したものである。
どの民族集団であっても、南アジア集団だけに共有されるハプロタイプ(青)、東南アジア集団だけに共有されるハプロタイプ(紫)、南アジアと東南アジアの両方の集団で共有されるハプロタイプ(緑)をあわせて90%以上となる。
はっきりとわかるのは、90%以上のハプロタイプが南アジア集団(青プラス緑)または東南アジア集団(緑プラス紫)と共有されていることである。
しかも東南アジアのみと共有する割合(緑)は、日本、韓国、華北、北方少数民族、ヤクートと北に行くほど低くなっている。
反対に、南アジアとも東南アジア集団とも共有しない常染色体SNPハプロタイプ(赤)の割合はたいへん低い。これらのハプロタイプこそ、中央アジアあるいはもっと北のルートを通って東アジアに来たことの証しだろう。
日本、韓国で2.8%、残りの東アジア全体(オロチョン、ダウール、ナナイ、蒙古、シベ、トゥ、イ、ナシ(Oroqen、Daur、Hazhen、Mongola、Xibo、Tu、Yi、Naxi))で5.6%と、共有群の多さ(90%以上)と好対照をなしている。
東アジアの北に分布するオロチョン、ダウール、ナナイ、蒙古、シベ、トゥ、イ、ナシ(残りの東アジア集団)は、南アジアや東南アジアと共有しないハプロタイプ(赤)が他の集団に比べて多いのだが、それでも南アジアや東南アジアと共有するハプロタイプの方が圧倒的に多いのである。
マクロで見たとき、南アジアから東南アジアを経て東アジアへ、すなわち南方ルートをとり、そこから南から北という方向に人が流れたことを示唆している。常染色体遺伝子でみる限り、中央アジア経由の割合は非常に限定的であったと言える。
2009年のサイエンス誌の論文(The HUGO Pan-Asian SNP Consortium 2009)は、仮説(A)を支持しているのである。
ⅲ)YdnaハプロタイプOの分布を再検証する
東アジア全体を特徴付けるハプロタイプOが仮説(A)に従って展開したという考えは妥当なのだろうか。すなわち、O集団が南方ルートで東南アジアに行き、そこから北に展開したという説にのっとり、O集団亜型それぞれの現在の分布を再検証してみよう。
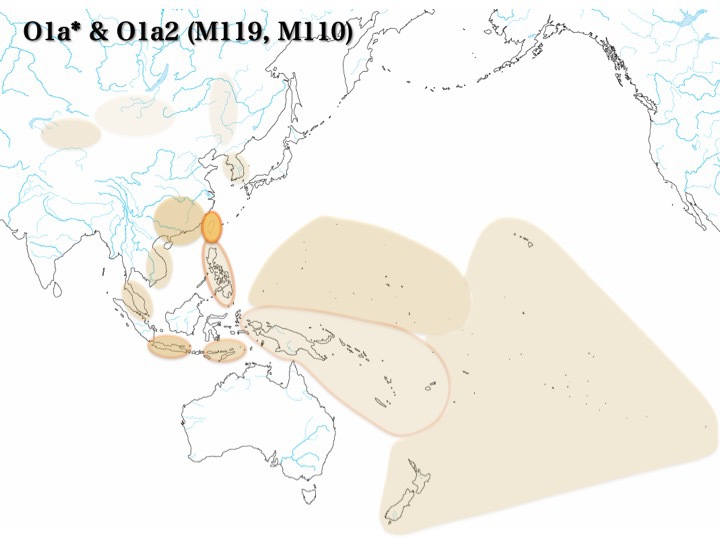
O1集団の分布
O1a*:塗りつぶし、O1a2:囲み線
塗りつぶし、囲み線、それぞれの濃度は集団の分布頻度を表す
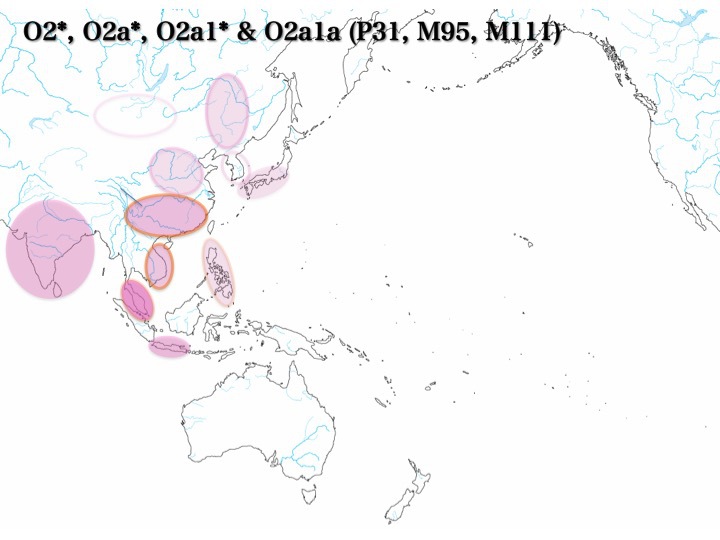
O2a集団の分布
O2*、O2a*:囲み線(ピンク色)、O2a1:塗りつぶし、O2a1a:囲み線(オレンジ色)
塗りつぶし、囲み線、それぞれの濃度は集団の分布頻度を表す
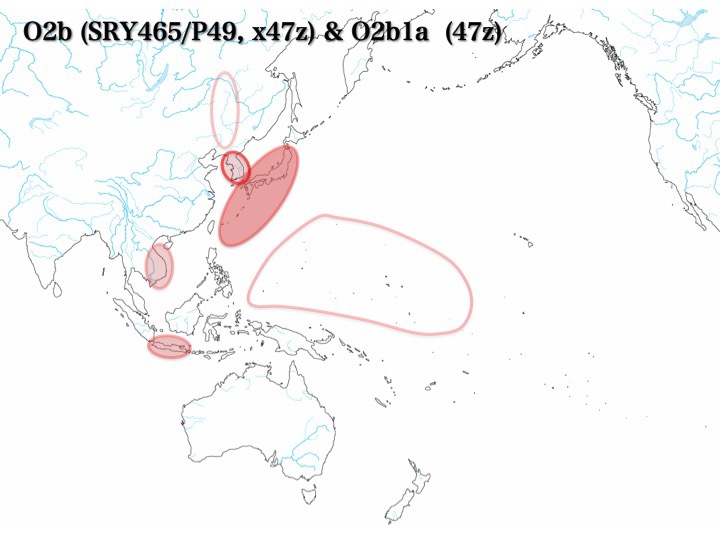
O2b集団の分布
O2b:囲み線、O2b1a:塗りつぶし
塗りつぶし、囲み線、それぞれの濃度は集団の分布頻度を表す
O3a1c集団の分布
塗りつぶしの濃度は分布頻度を表す
O3a2集団(上)、O3a2b集団(下)の分布
塗りつぶしの濃度は分布頻度を表す
O3a2c1集団の分布
塗りつぶしの濃度は分布頻度を表す
上記のように、O1、O2a、O2b、O3a1c、O3a2、O3a2b、およびO3a2cのどの集団も東南アジアには必ず分布している。
最北でも東南アジアの北に位置するベトナム、フィリピンに認められる。南ではマレー半島、インドネシアに存在する。各集団が南から北へ移動していったと想定しても不自然ではない。
ⅳ)ハプログループNOの発祥とNとOへの分岐、拡散
こうした点から眺めると、ハプログループNOとその子孫N、Oの分岐、移動拡散経路について、全く別の光景が見えて来る。
ハプログループNOの発祥は中央アジアとされた時期もあった。N集団はユーラシア大陸の北方に向かい、O集団は東アジアに行ったというものだ。しかし上述したような他の事実も付き合わせると、次にように考えるのが合理的なようだ。
すなわち、NOの発祥もNとOへの分岐も東南アジアで起こった。それぞれ北を目指して旅立った。Nはユーラシア大陸北端に東西にわたって幅広く分布した。O集団は東アジアにとどまって展開し、一部反転するように中央アジアに向かった。そう考えるのが合理的なようだ。
Ⅳ)日本人は南から来た
以上から何が言えるだろうか。ハプログループOはスペンサー・ウェルズのモデルのようにパミール高原、チベットを経てシナ大陸に入り、華南を経て東南アジアに向かったり、華北、華中、華南または朝鮮半島を経て日本に来たのだろうか。
スペンサー・ウェルズのモデルか
東南アジアから東アジア(南→北)方向の移動モデルか
(A)YdnaハプログループOの分布(左上図)Wikipediaによる(CC BY-SA 3.0)
(B)かつて考えられていたYdnaハプログループOの移動経路(左下図)「National Geographic Society の Genographic Project」による
(C)常染色体SNP(Single Nucleotide Polymorphism:一塩基多型)の大規模データ解析から得られた南→北ルートの可能性(右上図)。The HUGO Pan-Asian SNP Consortium 2009 より。
(D)YdnaハプログループOの海岸線沿い移動モデル(右下図)。
常染色体SNPの多様性を解析した結果、マクロの傾向として、出アフリカを果たした後で南アジアを経て東南アジアに入り、そこから北上した傾向がうかがえた(図右上)。Y染色体DNAハプログループO集団だけが、その大きな流れには乗らずに図左下の中央アジアルートを通ったと考えるのは不自然だろう。
それよりむしろC、Dと同様にYdnaハプログループO集団も、海岸線沿いのルートでスンダランドを含む東南アジアに至り、そこから北に転じてシナ大陸に入ったり、日本や朝鮮半島に移動定住したと考えられる(上図右下)。
先に述べたように、本稿では、日本人に特徴的なC、D、Oの各集団が最終氷期極期に入った頃から海水準が急激に上昇するまでの間、今から約30000~15000年前頃に日本列島に入って来たとする立場をとる。第一陣としてC、Dが、第二陣としてO集団が到来したというモデルだ。
こうした日本への到来モデルは「日本人のルーツは東南アジアにある」と言い換えることができる。次項では、この説を言語的な側面から検証しよう。
本稿の文献はこちらを参照されたい。
 Soliloquy1...
Soliloquy1...